


Plantas Daninhas? Serviços ecossistêmicos e a comunidade de espontâneas: Influência dos elementos da paisagem e da Intensidade de Manejo
índice
- 1. RESUMO
- 2. INTRODUÇÃO
- 3. REVISÃO DE LITERATURA
- 3.1 Paisagens Agrícolas Multifuncionais
- 3.2 Agrobiodiversidade
- 3.3 Plantas Espontâneas em Agroecossistemas como Elementos Geradores de Serviços, Diversificação e Usos Econômicos
- 3.4 Biodiversidade para os Agroecossistemas: Resiliência e Serviços Ecossistêmicos
- 4. MATERIAL E MÉTODOS
- 4.1 Caracterização da Área de Estudo
- 4.2 Caracterização da Amostragem Realizada
- 4.3 Métricas da Paisagem e de Intensidade de Manejo
- 4.3.1 Parâmetros avaliados na escala do cultivo (0,5 ha)
- 4.3.2 Parâmetros avaliados na escala da unidade produtiva (4,0 ha)
- 4.3.3 Parâmetros avaliados na escala da paisagem (95 ha)
- 4.4 Caracterização da Fertilidade do Solo
- 4.5 Caracterização da Comunidade de Espontâneas e do Banco de Sementes
- 4.6 Dinâmica de Condução do Experimento
- 4.7 Composição Fitossociológica e Distribuição das Espécies em Grupos Funcionais Prestadores de Serviços ao Agroecossistema
- 4.8 Análise Estatística
- 5. RESULTADOS E DISCUSSÃO
- 5.1 Métricas das Diferentes Escalas da Paisagem e de Intensidade de Manejo
- 5.1.1 Escala do cultivo (0,5 ha)
- 5.1.2 Escala da unidade produtiva (4,0 ha)
- 5.1.3 Escala da paisagem (95,0 ha)
- 5.2 Avaliação da Fertilidade do Solo
- 5.3 Descrição Geral da Comunidade de Espontâneas Amostradas in loco
- 5.4 Comunidade de Plantas Espontâneas do Banco de Sementes
- 5.4.1 Análise de agrupamento de espécies (cluster)
- 5.4.2 Agrupamento das espécies espontâneas pelo NMDS
- 5.5 Composição de Espontâneas em Grupos Funcionais e seus Serviços Ecossistêmicos Associados
- 5.6 Análise de Redundância (RDA) para a Associação dos Serviços Ecossistêmicos à Intensidade de Manejo e aos Componentes da Paisagem nas Escalas Avaliadas
- 6. CONCLUSÕES
- 7. CONSIDERAÇÕES FINAIS
- 8. REFERÊNCIAS BIBLIOGRÁFICAS
O texto publicado foi encaminhado por um usuário do site por meio do canal colaborativo Monografias. Brasil Escola não se responsabiliza pelo conteúdo do artigo publicado, que é de total responsabilidade do autor . Para acessar os textos produzidos pelo site, acesse: https://www.brasilescola.com.
1. RESUMO
Conciliar a produção de alimentos em quantidade e qualidade associada à conservação dos remanescentes florestais se aponta como uma das demandas emergenciais para o planeta, principalmente em ecossistemas tropicais. Pesquisas recentes vêm empenhando esforços para compreender e identificar grupos de espécies funcionalmente importantes para a manutenção da dinâmica dos agroecossistemas, e para o fornecimento de serviços ao ambiente. O presente trabalho teve como objetivo, mediante identificação dos padrões de práticas de manejo exercidas em sistemas agrícolas aliado aos dados de métrica da paisagem, compreender como diferentes vetores ambientais e de manejo influenciam a expressão de espécies e a composição funcional do banco de sementes de plantas espontâneas em diferentes escalas, avaliando seus possíveis efeitos no provimento de serviços ecossistêmicos proporcionados pelos diferentes grupos funcionais. O arranjo espacial do trabalho contempla 3 diferentes escalas, sendo: cultivo, unidade produtiva e paisagem, com as seguintes áreas: 0,5; 4,0 e 95 ha, respectivamente. O trabalho foi conduzido em duas glebas, distintas quanto à densidade de fragmentos florestais presentes, do assentamento São José da Boa Morte, no município de Cachoeiras de Macacu, RJ, Brasil. A caracterização das diferentes escalas se deu através de entrevistas semi-estruturadas junto aos agricultores e uso de ferramentas de geoprocessamento, onde foram obtidas informações sobre o manejo (i.e. uso de agrotóxico, tipo e intensidade de capina, ciclo da cultura e adubação do solo) e mensuradas algumas características ambientais (i.e. distância mínima linear cultivo-fragmento, áreas de vegetação secundária, monocultivos de arbóreas e frutíferas, fragmentos e contínuos florestais). Os resultados obtidos expressaram que práticas de manejo, principalmente a intensidade de capina, e a presença do elemento arbóreo na paisagem agrícola determinam a diversificação do banco de sementes de plantas espontâneas e a sua prestação de serviços ecossistêmicos. O trabalho elucida que o manejo da diversidade funcional de plantas espontâneas em agroecossistemas imersos em paisagens com pouca oferta de fontes de biodiversidade pode representar uma alternativa de baixo custo e aderente a realidade local, para o preenchimento de parte das lacunas funcionais existentes pela intensificação da agricultura.
Palavras-chave: Paisagens agrícolas, plantas espontâneas, banco de sementes, composição funcional, intensidade de manejo.
ABSTRACT
Conciliating food production in quantity and quality associated with the conservation of forest fragments is considered one of the emergency demands for the planet, especially in tropical ecosystems. Recent researches have making efforts to understand and identify groups of functionally important species to maintain the dynamics of agro-ecosystems, and to provide services to the environment. This study aimed, through the identification of management practices standards practiced in agricultural systems combined with landscape metrics data, understand how different environmental vectors and management influence the expression of species and the functional composition of the seed bank of spontaneous plants at different scales, assessing their possible effects in the provision of ecosystem services provided by different functional groups. The spatial arrangement of work includes three different scales: cultivation, productive unit and landscape, with the following areas: 0.5; 4,0 and 95 ha, respectively. The work was conducted in two plots, distinct in regards to the density of the present forest fragments in the São José da Boa Morte settlement, in Cachoeiras de Macacu, RJ, Brazil. The characterization of the different scales was made through semi-structured interviews with the farmers and using geoprocessing tools, where were obtained information about the management (i.e. use of pesticides, type and intensity of weeding, crop cycle and fertilization of the soil) and measured some environmental characteristics (i.e. minimum linear distance cultivation-fragment, areas of secondary vegetation, monoculture of arboreal and fruit trees, forest fragments and continuous forests). The results obtained showed that management practices, especially the intensity of weeding, and the presence of the tree element in the agricultural landscape determine the diversification of the seed bank of weed and its provision of ecosystem services. The work elucidates that the management of functional diversity of weeds in agricultural ecosystems immersed in landscapes with few options of biodiversity sources may represent a low cost alternative and suitable to local conditions, for the filling of part of the functional gaps due to the intensification of agriculture.
Keywords: Agricultural landscapes, weeds, seed bank, functional composition, management intensity.
2. INTRODUÇÃO
A agricultura atualmente representa um vetor de degradação ambiental, com forte impacto na alteração do clima, na perda da biodiversidade, na degradação do solo e na diminuição da oferta de água doce. Grandes danos têm sido constatados tanto na expansão da fronteira agrícola, quando áreas de vegetação nativa são convertidas a lavoura, quanto na intensificação da produção, através de técnicas produtivas convencionais, baseadas no aporte de insumos químicos e mecanização (FOLLEY et al., 2011).
Polinizadores, recursos genéticos para a melhoria de culturas agrícolas, habitat para predadores naturais promotores do controle biológico, microorganismos benéficos, ciclagem eficiente de nutrientes, manutenção da fertilidade do solo e regulação do ciclo hidrológico, são exemplos de serviços que recebem alta pressão por conta da simplificação de paisagens anteriormente complexas. Sendo assim, abordagens ajustadas a compreensão das funções ecossistêmicas estão sendo intensamente debatidas em todo o mundo (DORÉ et al., 2011), uma vez que são peças-chave no estabelecimento de paisagens agrícolas multifuncionais. A constituição dessas paisagens têm por objetivo a conservação dos recursos naturais e a manutenção da produtividade dos sistemas agrícolas.
Na escala da paisagem, a conservação da diversidade precisa ser estudada em diferentes escalas considerando o papel das práticas de manejo, no sistema e na unidade produtiva, e suas associações à estrutura da paisagem onde estão inseridos. Tais práticas, quando menos intensivas e mais heterogêneas, não só impulsionam um aumento da composição funcional no sistema, como promovem uma maior resiliência frente às adversidades ambientais e econômicas, como escassez hídrica, ataque de pragas e doenças e oscilações de mercado (JACKSON et al., 2007).
Nesse contexto, entre os organismos prestadores de serviços ecossistêmicos, as plantas espontâneas são muito relevantes, entre outras, por auxiliar na permeabilidade de paisagens agrícolas. A vegetação espontânea constitui a base da cadeia alimentar para produtores primários, herbívoros, fornecem abrigo e zonas de amortecimento, hibernação e reprodução (ex. ovoposição) e demais oportunidades à fauna (BÀRBERI et al., 2010). São largamente utilizadas na alimentação humana em diferentes partes do mundo, e possuem grande importância como plantas medicinais, sendo cada vez mais observadas por conta de inúmeros compostos potenciais de serem sintetizados para geração de medicamentos. Tais plantas também influenciam nas características edáficas, por promoverem diferentes composições de sistemas radiculares, podendo contribuir para uma melhor estruturação e aeração do solo em áreas antropizadas (COSTA & MITJA, 2009).
Em um momento em que o Brasil é considerado o líder do ranking de utilização de agrotóxicos do mundo e onde os herbicidas são 61% de todos os produtos formulados (IBAMA, 2014), se faz emergencial a geração de informações que deem suporte na readequação de práticas agrícolas visando uma maior sinergia das unidades produtivas com a paisagem agrícola.
Dito isso, considerando a vegetação espontânea como um dos elementos-chave para a manutenção da agrobiodiversidade, o presente trabalho teve como objetivo compreender, sob uma leitura integrada de diferentes escalas, se as diferentes intensidades do manejo em sistemas agrícolas associada à variação dos atributos da paisagem afeta a riqueza de espécies e a composição funcional do banco de sementes de plantas espontâneas. As hipóteses construídas são: (1) A presença de fragmentos florestais e do elemento arbóreo, associados ao manejo adotado, determinam a composição funcional da comunidade de espécies espontâneas prestadoras de serviços ecossistêmicos à produção agrícola; e (2) O uso constante de herbicidas empobrece o banco de sementes do solo, determinando lacunas funcionais.
3. REVISÃO DE LITERATURA
3.1. Paisagens Agrícolas Multifuncionais
A maioria das paisagens tropicais é constituída por mosaicos de vegetação que compreendem tanto manchas de vegetação natural quanto uma grande variedade de sistemas antropizados (ESTRADA & COATES-ESTRADA, 2002), sejam sistemas produtivos, áreas edificadas para moradia e produção, entre outros, cuja intensidade de manejo e densidade de ocorrência na paisagem determinam os impactos sobre a biodiversidade e processos ecossistêmicos (SLADE et al., 2011)
Identificar quais atividades podem ser proveitosamente combinadas na mesma localidade e quais são mais bem realizadas espacialmente separadas é uma questão importante nos aspectos teóricos e práticos quanto aos diferentes tipos de uso do solo (ANGLENSEN & KAIMOWITZ, 2001), principalmente em áreas que conciliam na mesma paisagem, atividade agrícola e a presença de vegetação natural com foco para a conservação. Tais elementos têm sido pauta para uma larga discussão no meio científico, dividindo percepções quanto a melhor maneira de atender as demandas por alimento e para a conservação da biodiversidade.
A dicotomia das estratégias poupa-terra versus compartilha-terra (land-sparing e land-sharing, no inglês) existe, pois enquanto a primeira propõe que as terras sejam parte destinadas para a conservação e parte para a produção de alimentos, separadamente, a seguinte propõe que menos áreas sejam destinadas especificamente para a conservação, entretanto em detrimento da redução da intensidade produtiva, objetivando manter a biodiversidade em toda a paisagem agrícola (GREEN et al., 2005). Esta última vem ganhando maior popularidade, sob o pressuposto de que a conservação é localmente compatível com a agricultura, particularmente em áreas tradicionais que executam manejo diversificado para múltiplos usos. Tais estratégias não são mutuamente excludentes, e muitos conservacionistas acreditam que uma combinação de ambos, valorizando a multifuncionalidade da paisagem, seja ideal para fornecer os vetores necessários para a conservação dos habitats naturais sem reduzir a oferta de produtos agrícolas (FISCHER et al., 2008; SCARIOT, 2013).
Por um lado, a multifuncionalidade é usada para caracterizar as atividades no setor da produção primária e do uso do solo, refletindo as consequências materiais das diversas demandas estabelecidas pela sociedade nos diferentes territórios, sendo então uma abordagem que se relaciona com a compreensão da multifuncionalidade agrícola. Por outro lado, esta também é utilizada para caracterizar a paisagem por si só.
A produção primária (i.e. agricultura, silvicultura, etc) é considerada função principal, incluindo uma mistura de bens materiais e imateriais, bem como a relação de bens públicos e privados (externalidades). Os produtos primários (ex. alimentos e fibras) não são os únicos serviços prestados em sistemas agrícolas, sendo o sequestro de CO2 e a recarga do lençol freático exemplos de serviços intangíveis.
Em sistemas naturais e agrícolas, particularmente, há a existência de um conjunto de espécies distintas quanto a sua abundância, composição e função (LOPES et al., 2011). Sendo assim, a tomada de decisões que impulsionem a conservação da diversidade local em paisagens multifuncionais, requer um conhecimento preciso e reprodutível sobre a composição das espécies em escala espacial e temporal, a fim de evitar equívocos em relação ao que tange a espacialidade e ao que diz respeito às influências do ambiente (GABRIEL et al., 2010; KORASAKI, 2013).
Espécies com diferentes características podem diferir em sua suscetibilidade ao isolamento e manejo das áreas (EWERS & DIDHAM, 2006), por exemplo, os predadores e parasitas de níveis tróficos superiores podem sofrer mais com o isolamento que seus hospedeiros ou presas (TSCHARNTKE et al., 2008; BAILEY et al., 2010).
Sendo assim, a incorporação de análises em múltiplas escalas no estudo de paisagens agrícolas tem sido observada como determinante para a compreensão da hierarquização de influência das diferentes escalas sobre a diversidade funcional de elementos bióticos (PETCHEY & GASTON, 2002; FAHRIG, 2013). Neste tocante, Gabriel et al. (2010) destaca dois pontos importantes: 1o) Grupos de espécies variam em suas exigências ecológicas, e por conta disso respondem de maneira diferente ao manejo exercido; 2o) A maioria dos estudos têm investigado o impacto da agricultura sobre a biodiversidade nas escalas de cultivo, mas as populações de muitos organismos respondem ao ambiente em escalas espaciais maiores. Ou seja, a maior parte destes estudos foi baseada em um modelo binário de paisagem ‘matriz-habitat’, que apresenta fortes limitações para muitos sistemas, já que paisagens reais são geralmente heterogêneas e muito mais complexas do que uma representação com apenas duas classes de cobertura do solo (MOREIRA et al., 2015).
Analisar a paisagem de maneira a integrar os diferentes componentes existentes, compreendendo a existência de relações em diferentes escalas, pode ser fundamental para a conservação e restauração de elementos benéficos ao sistema, como por exemplo, o carbono orgânico e a estrutura do solo, como para a conservação e uso sustentável dos recursos genéticos, promovendo a sensibilização do público para a importância da agrobiodiversidade e a sua relação com o avanço da segurança e soberania alimentar (BRUSSAARD et al., 2010).
3.2. Agrobiodiversidade
Resultante da interação entre o ambiente, recursos genéticos e práticas de manejo utilizadas pelas populações culturalmente diversas, a agrobiodiversidade varia em função das diferentes formas de uso do solo e engloba a variedade e diversidade de animais, plantas e microorganismos que são necessários para sustentar as funções chave, a estrutura e os processos do agroecossistema, oferecendo suporte para a produção de alimentos e segurança e soberania alimentar (VANDERMEER & PERFECTO, 1995; JACKSON et al., 2007). Portanto, o conhecimento e a cultura local devem ser considerados partes integrantes deste sistema, já que a atividade humana é determinante para a conservação (FAO, 1999a).
Ao longo de uma hierarquização ecológica, a agrobiodiversidade pode ser dividida em: (1) características genéticas e populacionais (ex. variedades tradicionais e espécies selvagens adaptadas às adversidades ambientais); (2) organismos que influenciam na produção agrícola e pecuária (ex. que reduzem a necessidade de insumos, agrotóxicos e outras externalidades); (3) heterogeneidade da biota em relação aos processos biofísicos dentro dos ecossistemas (ex. ciclagem de nutrientes derivada a partir da variação temporal e espacial da biodiversidade); e (4) interações na escala da paisagem entre o ecossistema agrícola e “não-agrícola” que melhoram os recursos na escala de cultivo, e potencialmente, a resiliência às adversidades ambientais (JACKSON et al., 2007).
As funções da agrobiodiversidade são mais bem compreendidas quando relacionadas à provisão de serviços como alimentos, combustível, fibra e produção de água doce do que para suporte (ex. ciclagem de nutrientes e formação do solo) e regulação (ex. clima, inundações, pragas e doenças ou purificação da água) (PASCUAL & PERRINGS, 2007).
Práticas convencionais de produção, tais como o uso intensivo de agrotóxicos e fertilizantes minerais, sistemas produtivos homogêneos, baseados em poucas variedades de alto rendimento, constante mecanização da área de cultivo através do uso de implementos pesados e remoção de habitats naturais e semi-naturais, tem resultado na simplificação das paisagens agrícolas em várias escalas espaciais (BENTON et al., 2003; HENDRICKX et al., 2007; PERFECTO & VANDERMEER, 2010).
Contudo, a erosão da biodiversidade tem sido uma das consequências das práticas agrícolas convencionais em todo o mundo e uma redução dos serviços ecossistêmicos já vêm sendo sentida. Há, portanto, uma crescente preocupação de que o declínio da biodiversidade comprometa o fornecimento dos serviços ecossistêmicos, comprometendo a produção agrícola e a vida humana (MEA, 2005). O controle biológico de pragas e doenças, provido por predadores e parasitóides tende a diminuir em paisagens intensivamente antropizadas (TSCHARNTKE et al., 2005; ALVES, 2013; ZHAO et al., 2015). A antropização e intensificação agrícola no uso de insumos químicos e maquinário também exerce efeitos negativos sobre a comunidade da fauna do solo e podem comprometer os serviços como a retenção de nutrientes, estruturação do solo (BRUSSAARD et al., 2007) e a ciclagem da matéria orgânica (JONGMANS et al., 2003).
O grande impacto negativo sobre o estoque milenar de agrobiodiversidade aconteceu a partir das políticas de modernização da agricultura, implementadas em diversas regiões do planeta a partir de meados do século XIX, sobretudo no século XX. Estimativas indicam que por conta das alterações abruptas dos ecossistemas naturais e agrícolas, no último século se perdeu de 50% a 90% da diversidade agrícola então existente (MULVANY & BERGER, 2003).
O Brasil é considerado um país megadiverso, detentor da maior biodiversidade do mundo (Conservation Internacional, 1998), sendo também berço natural de grande diversidade de plantas cultivadas. Este patrimônio genético e cultural constitui a base alimentar e a fonte de matéria-prima para inúmeras populações locais, além de ser elemento organizador de inúmeras expressões culturais. Conservá-los, portanto, é tarefa fundamental para a soberania alimentar destas populações e para a preservação do patrimônio cultural (OLIVEIRA et al., 2006).
Para neutralizar os efeitos negativos da intensificação da agricultura, devem-se expandir abordagens que conciliem a produção agrícola com a conservação e uso sustentável da biodiversidade e seus serviços ecossistêmicos associados (SCHERR & MCNEELY, 2008; BRUSSAARD et al., 2010), integrando agroecossistemas e buscando a compreensão da funcionalidade de diversos elementos provedores de serviços ao ambiente, traduzindo tal conhecimento em práticas agroecológicas e de manejo de paisagem, como a diversificação e rotação de culturas, adequada integração das áreas de cultivo com a vegetação natural, bordaduras, cercas-vivas e pequenos bosques em paisagens agrícolas (BIANCHI et al., 2013).
3.3. Plantas Espontâneas em Agroecossistemas como Elementos Geradores de Serviços, Diversificação e Usos Econômicos
Associadas a impactos negativos em agroecossistemas, principalmente baixos índices produtivos, plantas espontâneas têm sido comumente consideradas um grande problema para a agricultura, já que muitas espécies são reconhecidamente competidoras e/ou promovem alelopatia (DALE & POLASKY, 2007; ZHANG et al., 2007), havendo assim grandes esforços sendo realizados para promover melhores índices de controle e erradicação. Entretanto, diversas práticas tradicionais de manejo (ex. rotação de culturas) tendem a manter uma baixa densidade de espontâneas, porém não as erradicam do sistema.
Novas abordagens para o estudo da comunidade de espontâneas têm destacado os benefícios obtidos a partir destas plantas para o ambiente e ao bem-estar humano (MARSHALL et al., 2003; STORKEY et al., 2012). Diversas pesquisas têm oferecido informações quanto aos diferentes usos, serviços e funções desse tipo de vegetação para o agroecossistema, tais como o fornecimento de alimento para diferentes grupos tróficos, sequestro de carbono e estabilidade do solo (DALE & POLASKY, 2007; POWER, 2010). Alguns artrópodes fitófagos ao se alimentarem destas, são também beneficiados (BARBERI et al., 2010), como também várias aves e pequenos mamíferos (HOLLAND et al., 2006; ORLOWSKI & CZARNECKA, 2007; EVANS et al., 2011), regulando assim um sistema biótico que contribui para o controle biológico, reduzindo custos de produção.
Determinados organismos são diretamente afetados pelas mudanças na comunidade de espontâneas, como é o caso de insetos polinizadores, o que afeta a produção de algumas espécies vegetais (GABRIEL & TSCHARNTKE, 2007). É afetada também a biota do solo, por estarem intimamente relacionadas ao sistema radicular e a cobertura de copas, mas principalmente por conta do banco de sementes (FRANKE et al., 2009), que tem um papel fundamental na interação complexa destes organismos, servindo como alimento para larvas e nematoides benéficos. No entanto, as sementes também podem atuar como reservatórios para agentes patogênicos e vetores de doenças de plantas, podendo estes serem controlados com as práticas de rotação de culturas adequadas (FRANKE et al., 2009).
A variabilidade e densidade botânica de um banco de sementes em um determinado momento são o resultado de um balanço entre a entrada de novas sementes (por chuva e dispersão) e perdas por germinação, deterioração, parasitismo, predação e transporte (MACHADO et al., 2013). Segundo Carmona (1995), as semelhanças dos bancos de sementes entre os agroecossistemas são maiores para áreas mais intensificadas, tendo estas uma maior quantidade de propágulos em comparação a áreas que sofrem menor intensificação, pois plantas espontâneas possuem estratégias de alta produção de sementes em situações de maior estresse ambiental.
Práticas de manejo que promovam o revolvimento do solo como a aração e gradagem, podem estimular um aumento da viabilidade de propágulos, propiciando uma melhor distribuição de sementes dentro do perfil do solo, e também o aprofundamento de uma quantidade significativa destas sementes, reduzindo sua capacidade de regeneração. Por outro lado, as práticas que não revolvem camadas de solo podem permitir que a maioria das sementes permaneçam na superfície deste (LACERDA et al., 2005).
Fatores como a dormência e suas mudanças sazonais estão relacionados com a persistência de sementes no solo e, consequentemente, aos problemas de infestação em sistemas agrícolas, já que as sementes de espontâneas passam por ciclos anuais de maior ou menor intensidade de dormência. Estas mudanças são atribuídas a variações de temperatura, luz, chuvas, profundidade de sementes e as práticas agrícolas (VIVIAN et al., 2008).
O manejo da fertilidade do solo pode ser também considerado um importante componente associado à composição de plantas espontâneas (DI TOMASO, 1995; BLACKSHAW et al., 2004), pois a alteração da fertilidade afeta a densidade de espécies, sua dinâmica de absorção de nutrientes e produção de biomassa, impactando diretamente na biodiversidade (BLACKSHAW et al., 2005; MURPHY & LAMERLE, 2006). Por exemplo, Mahn et al. (1988) observou um declínio geral no número de indivíduos de espontâneas e um acréscimo de biomassa destas a partir do incremento de adubações nitrogenadas. Também afeta-se a relação entre as comunidades de Magnoliopsida/Liliopsida, que pode aumentar com a melhoria da fertilidade do solo (MACLEAN, 2003).
Tang et al. (2014) encontraram maior densidade de espontâneas em tratamentos contendo adubação nitrogenada e potássica em conjunto, tendo Yin et al. (2005) e Nie et al. (2008) reportado resultados similares para esta relação. Para os autores citados, a variabilidade estrutural da comunidade de espontâneas pode, entre outras, estar diretamente relacionada às diferentes condições de fertilidade do solo.
No entanto, poucos trabalhos até então se dedicaram a examinar o efeito da adubação na composição de plantas espontâneas em agroecossistemas (TANG et al., 2014), sendo importante o preenchimento dessa lacuna, já que a comunidade de espontâneas pode ser um elemento prestador de serviços ecossistêmicos. Sendo assim, fomentar a biodiversidade ao nível da escala de cultivo, mesmo impopular entre uma boa parcela dos agricultores devido ao risco de redução da produtividade por conta de competição com espécies espontâneas, se faz necessária para coincidir a produção agrícola com a conservação dos recursos biológicos, promovendo sistemas mais sustentáveis (STORKEY & CUSSANS, 2007; TANG, 2014).
Como ferramenta-chave do modelo de produção agrícola convencional, o uso de herbicidas continua a ser um dos principais motores de perda de biodiversidade em agroecossistemas (ULBER et al., 2009; ARMENGOT et al., 2013) e as altas diferenças encontradas na diversidade de espontâneas em áreas com sistema convencional em relação aos sistemas orgânicos são principalmente devido ao uso desses compostos. O uso de herbicidas pode promover o aparecimento de clones resistentes de plantas espontâneas generalistas, por exemplo, ao amplo-espectro não seletivo do glifosato (ANDREASEN & STREIBIG, 2011), pois repetidas aplicações de um mesmo herbicida ou herbicidas de mesmo mecanismo de ação podem causar uma pressão de seleção em indivíduos resistentes, que ocorrem em baixa frequência, tornando-os dominantes na população (CHRISTOFFOLETI et al., 2008).
Herbicidas diferem em termos de ingredientes ativos e modo de ação. Além disso, as espécies de plantas espontâneas também variam em suscetibilidade a esses produtos. A aplicação de herbicidas sintéticos de amplo-espectro em sistemas de cultivo convencionais podem exercer efeitos importantes sobre a riqueza dessa vegetação, sendo intensificado este reflexo ao se realizarem aplicações sucessivas, onde se altera tanto a estrutura de comunidades acima do solo como a composição do banco de sementes do mesmo (BÀRBERI et al., 1997).
O uso de herbicidas seletivos de espectro-estreito são intrinsecamente menos eficazes contra algumas espécies desejáveis de espontâneas de folha larga, portanto, tem sido apresentado como uma importante técnica de manejo dentro da produção, destinados a aumentar os benefícios ecológicos fornecidos por espécies da comunidade de espontâneas (SMITH et al., 2009). Consequentemente, controlando seletivamente espécies altamente competitivas, um equilíbrio pode ser alcançado entre um nível satisfatório de produtividade dos cultivos, sendo então importante que o adequado controle da vegetação espontânea atenda a necessidade de algumas espécies em apoiar a diversidade nos níveis tróficos superiores (ULBER, 2010).
Atualmente, novos enfoques quanto às aplicabilidades econômicas de plantas espontâneas vêm surgindo, principalmente pelo resgate do conhecimento tradicional em função de uma demanda potencial de consumo que encontra nesse estrato herbáceo, entre outras, a possibilidade de complementação alimentar, diversificação de cardápio e fonte de renda familiar. Sobretudo, nos países tropicais e subtropicais, a biodiversidade tem um grande potencial de uso alimentício a ser pesquisado (KINUPP, 2007).
Sistemas agrícolas imersos em paisagens complexas têm sido relatados como favoráveis à diversificação da comunidade de plantas espontâneas em comparação aos imersos em paisagens mais simplificadas, uma vez que o primeiro oferece um cenário em que existe uma maior quantidade de habitats, atuando como fonte de propágulos para o enriquecimento e manutenção do banco de sementes em áreas agrícolas (ARMENGOT et al., 2011). Desta forma, o banco de sementes pode refletir os efeitos da intensificação dos sistemas agrícolas ao longo do tempo, e por conta disso, prévios estudos têm reportado mudanças na riqueza e composição do banco de sementes de espécies espontâneas devido as diferentes intensidades de uso do solo e complexidade da paisagem agrícola (HAWES et al., 2010). Os resultados expressados por Gabriel et al. (2005) sinalizaram que no contexto avaliado, dentre todos os parâmetros o que melhor explicou a diversidade de plantas espontâneas foi a escala da paisagem (2 km de diâmetro), sugerindo que os padrões locais foram altamente determinados pelos atributos existentes em uma maior escala de abrangência. Entretanto o mesmo autor infere que a chuva de sementes de uma elevada diversidade de habitats e a existência de bordas perturbadas pode ser o processo subjacente mais provável para os dados obtidos, embora o mesmo sinalize que os mecanismos são em grande parte desconhecidos, sendo necessário o estímulo a pesquisas que contribuam na compreensão de tais interações, dando maior embasamento nas tomadas de decisão referentes ao manejo do agroecossistema utilizando a composição de espontâneas presente ou a potencial, existente no banco de sementes do solo.
3.4. Biodiversidade para os Agroecossistemas: Resiliência e Serviços Ecossistêmicos
Em paisagens complexas, a biodiversidade tende a ser elevada, o que significa existir um risco substancial de perda de elementos funcionais quando há uma matriz intensificada. Esta mesma matriz pode receber um alto nível de imigração dos organismos vindos de fragmentos florestais, podendo compensar alguns efeitos deletérios das práticas agrícolas locais (BIANCHI et al., 2006; RICKETTS et al., 2008).
No sentido de compreender melhor como os processos de intensificação da agricultura têm afetado a diversidade de espécies e a provisão de seus serviços ecossistêmicos associados, pesquisas recentes vêm se mostrando atentas às consequências em nível de diversidade funcional (i. e. valor e alcance de características funcionais dos organismos em uma comunidade) em diferentes escalas dos agroecossistemas (HOOPER et al., 2005).
Supõe-se geralmente que a perda de espécies em função do uso intensivo do solo resulta em uma similar perda da diversidade funcional, no entanto, esta suposição vem sendo questionada (MAYFIELD et al., 2010). Embora frequentemente haja uma positiva correlação quando feita esta avaliação na escala de cultivo (ARMENGOT et al., 2011; STORKEY et al., 2012; GUERRERO et al., 2010, 2014), pode-se ocorrer cursos diferentes a depender do grau de redundância funcional na comunidade (i. e. o número de espécies possuindo características funcionais semelhantes) e como as mudanças no manejo da paisagem afetam a organização dos processos da comunidade. De acordo com Flynn et al. (2009), a riqueza de espécies pode declinar sem que haja uma perda correspondente da diversidade funcional em comunidades com alta redundância funcional e pode ser até mesmo possível o aumento da diversidade funcional sem uma mudança na riqueza de espécies, a depender das mudanças nos filtros ambientais, favorecendo o aparecimento de espécie funcionalmente diferentes.
Resiliência é a capacidade de um sistema em se organizar após uma adversidade, retendo essencialmente a mesma função, estrutura e identidade (PIMM, 1991). Ferer et al. (2002) destacam que a quantificação da resiliência é complexa e que a escolha de componentes ou propriedades do ecossistema que constituem os parâmetros que suportam a expressão quantitativa de resiliência devem considerar os diferentes níveis de organização dos sistemas ecológicos.
Uma das formas de ampliação da resiliência em agroecossistemas se dá através da diversificação de sistemas agrícolas através de práticas que incorporem a agrobiodiversidade em uma múltipla escala espacial e/ou temporal (ALTIERI, 2004; TOMICH et al., 2011; KREMEN et al., 2012). Sistemas agrícolas diversificados têm muito em comum com a abordagem multifuncional e agroecológica quanto ao seu modo de manejo e resultados. Os indicadores-chave em tais sistemas servem como mecanismos para a manutenção e regeneração de interações bióticas e, por sua vez, os serviços ecossistêmicos, como por exemplo, a qualidade do solo, a fixação de nitrogênio, polinização e controle de pragas e doenças, fornecendo benefícios essenciais para a agricultura.
A prática de consórcio de culturas é pensado para gerar uma sobre-produção, pois diferentes plantas cultivadas em conjunto podem utilizar mais dos recursos disponíveis (ex. diferentes profundidades radiculares acessando uma maior fração de nutrientes espacialmente estratificados e água) ou por conta da facilitação que uma planta pode promover a outra (HAUGGARD-NEILSEN & JENSEN, 2005). Ao promover a diversidade floral através de sebes ou manutenção de áreas seminaturais, produtores podem melhorar a atração de inimigos naturais e/ou polinizadores para suas lavouras, aumentando o controle de pragas (KREMEN et al., 2002; MORANDIN & WINSTON, 2005, LETOURNEAU et al., 2011). O fortalecimento desses serviços ecossistêmicos tão somente é possível com a manutenção das práticas de cultivo diversificadas nas diferentes escalas, auxiliando na ampliação da sua resiliência frente às adversidades ambientais como períodos de estresse hídrico, excesso de chuva ou infestação de pragas e doenças (LIN, 2011).
Estudos em controle biológico que se atentam em averiguar a influência da paisagem circundante sobre a regulação de pragas (BIANCHI et al., 2006), recentemente têm apontado que a complexidade da paisagem (i. e. qualidade e quantidade de vegetação secundária em torno de unidades produtivas) pode afetar significativamente no controle da infestação de pragas (GARDINER et al., 2009). Uma recente meta-análise (CHAPLIN-KRAMER et al., 2011) que comparou locais em diferentes contextos da paisagem constatou que não houve efeitos significativos para as respostas da abundância de pragas e danos nas culturas agrícolas, entretanto, a abundância, diversidade e as taxas de predação ou parasitismo aumentou significativamente com a ampliação da complexidade da paisagem.
Assim, o uso de sistemas agrícolas diversificados possibilitam maior chance de serem contemplados com diferentes tipos de serviços ecossistêmicos, sendo potencial estratégia para aumentar a produção e a segurança alimentar local, particularmente em áreas consideradas hotspots de biodiversidade (CINCOTTA et al., 2000), que por sua vez poderia potencialmente reduzir a necessidade da conversão de habitat reduzindo a perda de biodiversidade (PHALAN et al., 2011).
4. MATERIAL E MÉTODOS
4.1. Caracterização da Área de Estudo
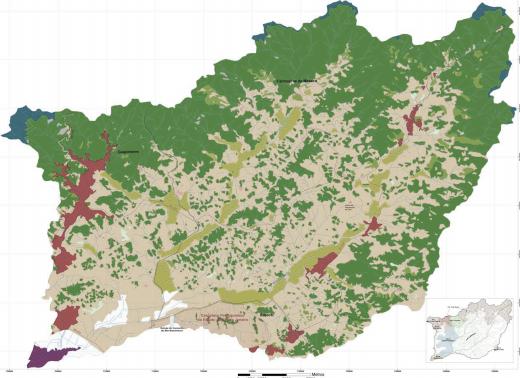
O estudo foi conduzido no assentamento rural São José da Boa Morte, localizado no município de Cachoeiras de Macacu, RJ, Brasil. O município é parte na Bacia Hidrográfica do Rio Guapi-Macacu (Fig. 1).
Fonte: Embrapa Solos
Figura 1. Esquema representativo do estado do Rio de Janeiro, da Bacia Guapi-Macacu e o assentamento São José da Boa Morte.
O clima é classificado como tropical úmido, inserido nas zonas climáticas Aw/Cfa de acordo com a classificação de Köppen-Geiger (KOTTEK et al., 2006). O período de estiagem na região ocorre de maio a setembro, sendo julho o mês mais seco com média de precipitação de 32 mm. O período de maior precipitação compreende os meses de novembro a março, com média de 208 mm em dezembro. A precipitação média acumulada anualmente é de 1307 mm. A temperatura média é de 23.1°C, sendo julho o mês mais frio, com mínimas de 14.8°C. Fevereiro é o mês mais quente, com máximas de 31°C (CLIMATE DATA, 2013). O assentamento, alvo deste estudo, tem como predominância solos das ordens dos Gleissolos, Latossolos e Organossolos.
De acordo com Fidalgo et al. (2008), as terras da Bacia são ocupadas predominantemente por florestas (45%) do tipo Ombrófila Densa. As unidades de conservação de proteção integral, representadas pelo Parque Estadual dos Três Picos, a Estação Ecológica Estadual do Paraíso e o Parque Nacional da Serra dos Órgãos, ocupam aproximadamente 25,6% da área da Bacia, e as unidades de conservação de uso sustentável, representadas pelas Áreas de Proteção Ambiental (APA) da Bacia do Rio Macacu, de Guapiaçu, de Petrópolis, de Guapimirim e do Rio São João – Mico Leão Dourado ocupam 37,2%. O assentamento São José da Boa Morte encontra-se dentro da APA da Bacia do Rio Guapi-Macacu.
O mesmo estudo aponta que áreas de pastagens representam 43,6% da Bacia, contrastando com os 4,8% referentes às áreas de produção agrícola, que estão concentradas ao longo de rodovias e rios principais, em locais de mais fácil acesso. Ainda que com percentual pouco representativo em comparação aos demais usos da terra, o município de Cachoeira de Macacu tem grande destaque agrícola no estado do Rio de Janeiro, uma vez que concentra as áreas de baixadas, onde são cultivadas olerícolas como inhame (Colocasia spp.), jiló (Solanum gilo) e quiabo (Hibiscus esculentus); feijão (Phaseolus vulgaris), milho (Zea mays) e aipim (Manihot spp.), e também, frutíferas como goiaba (Psidium guayava), maracujá (Passiflora edulis), laranja (Citrus spp.), coco-verde (Cocos nucifera) e banana (Musa spp.). A região representa uma resistência frente a um Estado declaradamente não-agrícola.
O município se encontra em 2º lugar no ranking dos repasses do ICMS-Verde para o Estado do Rio de Janeiro (SEA, 2014), entretanto, não é reconhecida a importância dos sistemas agrícolas na manutenção das áreas de vegetação natural e da biodiversidade da paisagem, o que redunda em uma estratégia de conservação que não considera a vocação agrícola do município (UZÊDA, 2012).
4.2. Caracterização da Amostragem Realizada
As glebas que subdividem o assentamento São José da Boa Morte possuem diferentes níveis de antropização, indo desde áreas densamente ocupadas por fragmentos florestais até as fortemente alteradas, onde a vegetação nativa é praticamente inexistente.
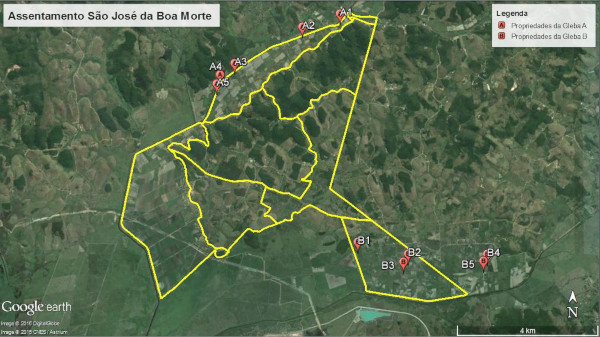
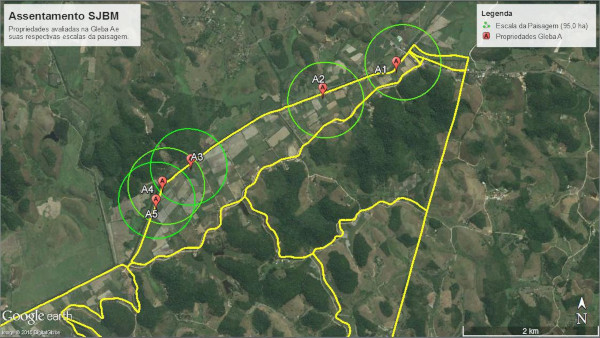
A escolha das unidades produtivas avaliadas se deu em função da sua localização dentro do gradiente de antropização encontrado na paisagem do assentamento, com o intuito de evidenciar a influência dos fragmentos florestais na composição da comunidade de plantas espontâneas. Sendo assim, foram selecionadas 5 unidades produtivas na “Gleba A”, área com alta densidade de fragmentos florestais e 5 unidades produtivas na “Gleba B”, fortemente desprovida de fragmentos de vegetação nativa (Fig. 2).
Em cada uma das glebas foram inclusos um sistema que apresentou características interessantes de serem observadas quanto ao possível impacto deste ao banco de sementes do solo, sendo o da “Gleba A” um sistema em que foi feito uso de adubação-verde com “mucuna” (Mucuna spp.) no ciclo anterior do cultivo existente e na “Gleba B” um sistema em que parte da área realizou-se capina seletiva e a outra parte o uso de herbicida, totalizando 6 sistemas em cada uma das glebas.
Fonte: Do próprio autor/Google Earth
Figura 2. Visão geral contendo as unidades produtivas na Gleba A e Gleba B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ.
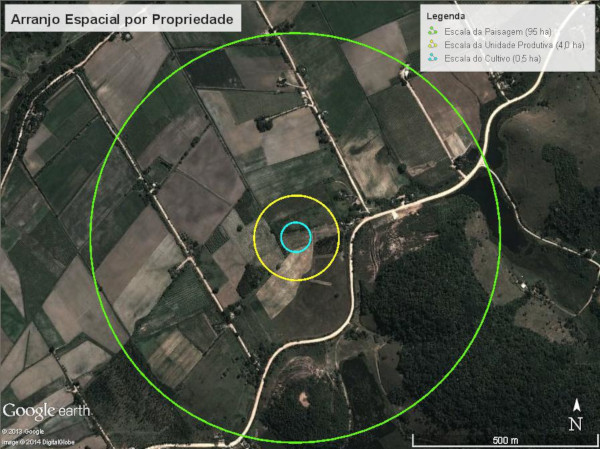
A fim de melhor compreender as interações existentes entre as diferentes escalas da paisagem, de forma a observar a influência desta interação na composição da comunidade de espontâneas, realizou-se o seguinte arranjo espacial (Fig. 3):
-
Escala do Cultivo: Área com a presença de uma cultura agrícola, tendo tamanho médio de 0,5 ha ou raio de 40 m.
-
Escala da Unidade Produtiva: Áreas com múltiplos cultivos, tendo tamanho médio de 4,0 ha ou raio de 110 m desde o centro do cultivo.
-
Escala da Paisagem: Conjuntos de diferentes usos do solo onde estão inseridas as propriedades e cultivos, tendo tamanho médio de 95 ha ou raio de 550 m desde o centro do cultivo.
Fonte: Do próprio autor/Google Earth
Figura 3. Detalhe das 3 escalas de análise: círculo menor: Cultivo (0,5 ha); círculo intermediário: Unidade Produtiva (4,0 ha) e círculo maior: Paisagem (95 ha), realizadas em cada propriedade avaliada nas glebas A e B do assentamento São José da Boa Morte, RJ.
4.3. Métricas da Paisagem e de Intensidade de Manejo
A caracterização do manejo da Escala do Cultivo (0,5 ha) e da Unidade Produtiva (4,0 ha) foi realizada através de entrevistas semi-estruturadas junto aos agricultores.
Foram levantadas informações quanto aos principais pontos relativos às culturas implantadas na propriedade, manejo do solo e utilização de insumos. A partir dos dados coletados construiu-se um ‘Índice de Qualidade de Manejo’, descrito a seguir.
4.3.1. Parâmetros avaliados na escala do cultivo (0,5 ha)
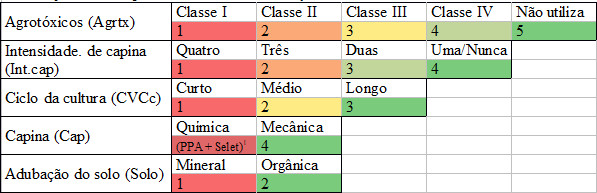
Visando identificar a influência que o manejo adotado nos diferente sistemas produtivos pode promover sobre o banco de sementes de plantas espontâneas, foram definidos parâmetros que exercem uma potencial pressão de seleção sobre a comunidade de espontâneas. São eles: Ciclo da cultura ; Tipo e número de capinas realizadas; Potencial de Periculosidade Ambiental (PPA) dos agrotóxicos utilizados; e Manejo da adubação do solo.
Para cada parâmetro avaliado foi estabelecido uma escala de impacto, com valor máximo e mínimo, em função das especificidades do mesmo, como mostra a Tabela 1.
Na escala apresentada na tabela a seguir os valores baixos representativos de um maior nível de impacto do determinado parâmetro ao sistema e os valores altos, o oposto.
Tabela 1. Parâmetros mensurados na escala do cultivo (0,5 ha) para avaliação da intensidade de manejo e suas respectivas escalas de valoração.
Fonte: Do próprio autor
1(PPA + Selet): Representa o cálculo para determinar o valor de capina química (uso de herbicida), sendo: PPA (1, 2, 3 ou 4)*(0,6) + Seletividade do herbicida (1 = Não-seletiva; 2 = Seletiva para dicotiledônea; 3 = Seletiva para monocotiledônea)*(0,4), sendo o valor final sempre menor que 4.
A avaliação do Potencial de Periculosidade Ambiental (PPA) é baseada em estudos laboratoriais que demonstram as características do produto, suas propriedades físico-químicas, sua toxidade a diversos níveis tróficos (microrganismos, minhocas, microcrustáceos, algas, peixes, aves, abelhas e mamíferos), bioacumulação, persistência (biodegradabilidade no solo, hidrólise e fotólise), bem como os potenciais mutagênicos, carcinogênicos e embriofetotóxicos.
O PPA é dividido em diferentes classes, que variam de I a IV, sendo: produtos altamente perigosos ao meio ambiente (Classe I, rótulo vermelho), produtos muito perigosos ao meio ambiente (Classe II, rótulo amarelo), produtos perigosos ao meio ambiente (Classe III, rótulo azul) e produtos pouco perigosos ao meio ambiente (Classe IV, rótulo verde).
A periculosidade ambiental é, portanto, atribuída as características do produto que promovem contaminação e danos aos compartimentos bióticos e abióticos dos ecossistemas (RIEDER, 2004), desta forma, esta classificação de fácil obtenção, pode ser utilizada como um dos componentes indicativos da intensidade de manejo de um sistema de produção agrícola.O parâmetro “Potencial de Periculosidade Ambiental (PPA) dos agrotóxicos utilizados” foi então utilizado para obter a classificação (via rotulagem) (Tabela 3) dos produtos comerciais utilizados na condução da cultura agrícola na época da realização do estudo. Os herbicidas levantados foram classificados posteriormente no parâmetro “Capina”. Pontuação varia de 1 a 5.
O parâmetro “Tipo e número de capinas realizadas” se faz determinante na compreensão e discussão dos resultados gerados neste trabalho, pois as diferentes práticas de controle da vegetação espontânea podem impactar com diferentes intensidades os atributos físicos, químicos e biológicos do solo (DALE & POLASKY, 2007). Sendo assim, buscou-se junto aos agricultores identificar qual o manejo realizado no controle de plantas espontâneas durante o ciclo da cultura agrícola presente no 0,5 ha (escala do cultivo) no momento do estudo.
De acordo com os relatos dos agricultores, escalonou-se a intensidade de capina (Int.cap.) em: Uma vez/Nenhuma; Duas vezes; Três vezes e Quatro vezes. A pontuação varia de 1 a 4, sendo considerado que quanto maior o número de capinas, mais negativo é o impacto aos componentes edáficos e por isso menor pontuação.
O ciclo de vida da cultura (CVC.c) foi definido em: Curto (≤ 4 meses); Médio (> 4 meses e ≤ 2 anos); e Perene (> 2 anos) (Tab. 2). Pontuação varia de 1 a 3.
Tabela 2. Culturas agrícolas presentes nas áreas de 0,5 ha (escala do cultivo) das glebas A e B do assentamento São José da Boa Morte (RJ) e ciclo de cultivo, março/2014.
|
Cultura |
Ciclo |
|
Batata-doce |
Curto |
|
Feijão-mauá |
Curto |
|
Jiló |
Curto |
|
Pimentão |
Curto |
|
Quiabo |
Curto |
|
Mandioca |
Médio |
|
Milho |
Médio |
|
Goiaba |
Perene |
Fonte: Do próprio autor
Quanto ao tipo de capina, pode ser classificada em: Mecânica (uso de enxada, tobata, roçadeira costal, microtrator, etc.) e Química (uso de herbicida sintético).
Consideramos nesse item que o controle mecânico tende a ser menos impactante na diversidade do banco de sementes que o controle químico. A pontuação para capina mecânica é 4 e para capina química é um somatório de um produto, sendo: PPA*(0,6) + Seletividade*(0,4), por considerarmos que os parâmetros avaliados na classificação do potencial de periculosidade ambiental devam receber uma ponderação maior que a seletividade.
O PPA foi valorado quanto a sua classificação padrão (1 a 4) e a seletividade varia de 1 a 3, respectivamente ao espectro de ação dos herbicidas: não-seletivo, seletivo a folha larga (dicotiledônea), seletivo a folha estreita (monocotiledôneas). Esta ordem reflete o que consideramos ser mais adequado no que tange as funcionalidades do estrato herbáceo, e neste caso, plantas que provém uma maior diversidade de flores podem conferir distintos benefícios à unidade produtiva, como por exemplo, a atração de inimigos naturais e polinizadores (TSCHARNTKE et al., 2007; CHAPLIN-KRAMER et al., 2013).
Para ‘Manejo da adubação do solo’, classificou-se em: Orgânica (uso de esterco animal, composto, farinha de osso, etc) e Mineral (uso de fertilizantes minerais). Pontuação varia de 1 a 2, sendo a adubação orgânica considerada a que promove melhor aporte de qualidade ao solo, desde que realizada corretamente.
No item a seguir serão descritas as métricas avaliadas na escala da unidade produtiva (4,0 ha), que envolveu os sistemas avaliados neste item.
4.3.2. Parâmetros avaliados na escala da unidade produtiva (4,0 ha)
A caracterização da unidade produtiva foi realizada por base na composição de cultivos existentes e na presença do elemento arbóreo agrupado ou disperso.
Esta avaliação se deu através da criação de croquis da unidade produtiva junto aos agricultores e utilização das ferramentas de métrica do software ArcGIS 10.2.1.
Parâmetros:
- Ciclo de Cultivo (CVC.up): Similar ao levantamento realizado na escala do cultivo, este parâmetro visa classificar as culturas agrícolas existentes na área de 4,0 ha quanto ao seu ciclo de cultivo (curto, médio e perene);
- Elemento Arbóreo (EA.up): Presença de cerca viva com uma ou várias espécies, vegetação de beira de rio, sendo espécies nativas ou não e áreas de vegetação secundária menor que 0,5 ha, sendo eles divididos em duas classes:
-
Monocultivo (EA.up 1): Áreas com coco, eucalipto e goiaba;
-
Elemento arbóreo (EA.up 2): Todo o restante.
- Área de Fragmentos Florestais (AFF.up): Áreas com vegetação nativa maiores que 0,5 ha;
4.3.3. Parâmetros avaliados na escala da paisagem (95 ha)
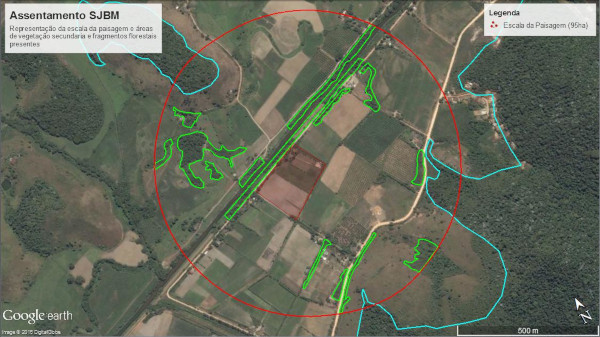
A escala da paisagem representa a área em que as fontes de biodiversidade podem estar contidas, a depender da complexidade da mesma (Fig. 4 e 5).
Fonte: Do próprio autor/Google Earth
Figura 4. Detalhe da Gleba A, destacando o as propriedades analisadas, a circunferência da escala da paisagem e a presença de fragmentos florestais de maior densidade.
.jpg)
Fonte: Do próprio autor/Google Earth
Figura 5. Detalhe da Gleba B, destacando as propriedades analisadas e a ausência de fragmentos florestais de maior porte.
Fonte: Do próprio autor/Google Earth
Figura 6. Recorte na escala da paisagem (95 ha), com destaque para as áreas de vegetação secundária e fragmentos florestais destacados em verde e azul, respectivamente.
Parâmetros:
Foram utilizados os mesmos parâmetros “Área de fragmentos florestais” (AFF.p) e “Elemento arbóreo” (EA.p), já caracterizados no item anterior (3.3.2 - Parâmetros avaliados na escala da unidade produtiva - 4,0 ha), sendo adicionado a esta avaliação:
- Distância cultivo-fragmento (Dist.p): Distância linear (m) do ponto focal (0,5 ha) ao fragmento florestal mais próximo;
- Área total do fragmento mais próximo (Área.frag): Mensuração da área total do fragmento mais próximo ao cultivo central (escala do cultivo - 0,5 ha).
4.4. Caracterização da Fertilidade do Solo
A fim de caracterizar os sistemas avaliados e observar se a fertilidade do solo influencia na composição funcional de espontâneas, foi realizada a amostragem do solo utilizando os mesmos pontos de coleta para as demais avaliações.
Realizou-se a amostragem nas profundidades 0-5 cm e 5-10 cm por ponto, utilizando um trado holandês.
As amostras devidamente identificadas foram secas em TFSA (terra fina seca ao ar) e encaminhadas ao Laboratório de Química Agrícola da Embrapa Agrobiologia para a avaliação total dos macroelementos do solo, sendo eles e seus respectivos métodos utilizados: ‘C’ – Walkey & Black; ‘Al’ – Titulação; ‘Ca’ – Absorção atômica; ‘K’ – Fotometria de chama; ‘Mg’ – Absorção atômica; ‘N’ – Kjedahl; ‘P’- Colorimétrico, além da acidez titulável (‘H+Al’ - Titulação) e do ‘pH’ - Potenciometria. Os procedimentos foram baseados no "Manual de Laboratórios: Solo, Água, Nutrição Animal e Alimentos – Embrapa, 2005".
4.5. Caracterização da Comunidade de Espontâneas e do Banco de Sementes
A amostragem e coleta da vegetação espontânea e dos monólitos de solo foram realizada nas áreas de 0,5 ha, referentes à escala do cultivo, onde foram feitas coletas em 5 pontos da área, em zig-zag e distantes aproximadamente 25 m entre si (Fig. 7). Cada ponto foi marcado em GPS.
As coletas aconteceram em dois momentos, sendo:
- Composição da Comunidade de Espontâneas:
Foram coletadas no mês de março de 2014, destinado a descrição geral da composição de espontâneas, sendo utilizado um gabarito de 0,25 m² (0,5 m x 0,5 m) para a amostragem. O material obtido em cada um dos 5 pontos foi, através de suas características morfológicas, separado em 3 grupos (Cyperáceas, Dicotiledôneas e Monocotiledôneas). A inclusão do grupo Cyperáceas se dá em função da importância que muitas das plantas pertencentes a família Cyperaceae possui quanto ao impacto de plantas espontâneas em agroecossistemas, sendo de interesse observar tal característica no contexto do presente trabalho, e motivando a sua separação do grupo que inclui as plantas da classe das monocotiledôneas. Posteriormente o material foi levado à estufa de circulação forçada onde permaneciam em temperatura média de 65ºC até atingirem peso constante. Ao final o material seco era pesado a fim de saber a contribuição em massa de cada grupo ao sistema.
- Banco de Sementes do Solo:
A caracterização da comunidade de espontâneas a partir do banco de sementes se deu a partir de monólitos de solo coletados nos sistemas de cultivo avaliados. Foram coletados 62 monólitos de solo nos 12 sistemas de cultivo avaliados.
Em cada cultivo foram coletados 10 monólitos de solo nas dimensões (18,0 x 13,5 x 9,0 cm), retirados com o auxilio de uma pá-reta para o corte do solo e uma faca. Em seguida foram envoltos em sacolas plásticas devidamente identificadas e acomodados em caixas de papelão tipo Sedex (Fig. 8) sendo posteriormente destorroado para facilitar a expressão da germinação do banco de sementes.

Fonte: Do próprio autor/Google Earth
Figura 7. Representação da distribuição dos pontos de coleta na área de 0,5 ha (escala do cultivo)

Fonte: Do próprio autor
Figura 8. Representação do material coletado devidamente acomodado para transporte.
4.6. Dinâmica de Condução do Experimento
O material coletado foi direcionado a uma casa de vegetação pertencente às instalações da Embrapa Agrobiologia, onde o experimento foi instalado e conduzido.
As amostras foram dispostas, individualmente, em bandejas perfuradas com dimensões (25 x 19,8 x 3,1 cm), devidamente identificadas e envoltas em sacolas plásticas de cor preta a fim de evitar o contato direto do material com o alumínio e às alterações de luz pela refletância do mesmo.
Para cada ponto amostrado foram dispostas duas bandejas, sendo uma com o monólito indeformado, a fim de observar a germinação das plântulas quando conservada as características do solo (i.e. compactação, estruturação, residual de herbicidas, espacialidade das sementes, entre outros) e a outra como um par deste monólito, porém fragmentado e deformado, a fim de eliminar filtros naturais e promover melhores condições para a expressão do banco de sementes (Fig. 9). Elucida-se que para este trabalho foram utilizadas como base as amostras deformadas.
As bandejas foram irrigadas diariamente, com turnos distintos em função das condições climáticas.
O método utilizado para a quantificação das plântulas foi o de emergência de plântulas ou germinação, adaptado de Gross (1990). A germinação foi acompanhada durante 14 semanas, tendo sido realizados quatro censos, mensalmente, entre julho e outubro de 2014.
O censo consistia em quantificar e separar indivíduos de mesmo morfótipo, alocando-os agrupados em sacos de papel devidamente identificados. Ao final de cada censo eram retirados todos os indivíduos de plântulas e realizava-se o revolvimento da terra.
Os indivíduos jovens, cuja identificação permaneceu duvidosa, foram transplantados para recipientes maiores com plaquetas de identificação, até desenvolverem-se a ponto de possibilitar a sua identificação a nível de espécie.
A identificação das espécies foi feita por consulta à bibliografia botânica, e por auxílio de especialistas do Herbário RBR (Departamento de Botânica/ IB/ UFRRJ).

Fonte: Do próprio autor
Figura 9. Disposição das bandejas para a metodologia de emergência de plântulas, sendo posicionadas à esquerda as amostras deformadas.
4.7. Composição Fitossociológica e Distribuição das Espécies em Grupos Funcionais Prestadores de Serviços ao Agroecossistema
Diversidade funcional é a extensão das diferentes funcionalidades entre as espécies em uma comunidade (TILMAN, 2001), sendo um importante determinante dos processos dos ecossistemas (LOREAU et al., 2001). Uma medida comum da diversidade funcional é o número de grupos funcionais representados pelas espécies de uma comunidade (PETCHEY & GASTON, 2002).
Realizou-se mediante busca em fontes bibliográficas consolidadas (ex. Science Direct) um levantamento quanto aos potenciais de uso e serviços ecossistêmicos das espécies de plantas espontâneas emergidas na avaliação da dinâmica do banco de sementes do solo.
Foram pré-determinados 4 grupos funcionais e seus serviços associados (Tab. 4).
Tabela 3. Grupos funcionais de plantas espontâneas e seus serviços ecossistêmicos associados
|
GRUPOS FUNCIONAIS |
SERVIÇOS ECOSSISTÊMICOS PRESTADOS |
|
Alimentação e Saúde Humana - (ASH) |
|
|
Aumento da Qualidade do Solo - (AQS) |
|
|
Controle de Pragas e Doenças - (CPD) |
|
|
Estímulo a Polinização - (EP) |
|
Fonte: Do próprio autor
Posteriormente ao levantamento das características funcionais por espécie, foi realizada a aglutinação nos diferentes grupos funcionais propostos, de todas as espécies emergidas do banco de sementes com o intuito de realizar inferências quanto aos principais serviços ecossistêmicos existentes em cada área de estudo e como o estrato herbáceo, representado pela vegetação espontânea, tem contribuído em conjunto no cumprimento destes serviços.
4.8. Análise Estatística
Para o cálculo dos índices referentes à análise da comunidade de espontâneas oriunda do banco de sementes (Abundâncias total e relativa, Riqueza de espécies, Frequência relativa, Diversidade de Shannon-Wiener) foram utilizadas as ferramentas de planilha dinâmica do software Microsoft Excel, presente no pacote Office 2010. Para análise dos demais dados foi utilizado o software R (R Development Core Team, 2004). Para avaliação da similaridade na composição da comunidade de espécies encontradas nas diferentes unidades produtivas foi utilizada a análise de agrupamento (cluster analysis) também denominado classificação não supervisionada, utilizando coeficientes de distância euclidiana pelo método de Bray-Curtis a partir dos dados de frequência. A análise de escalonamento multidimensional (NMDS, Nonmetric Multidimensional Scaling) teve como objetivo detectar dimensões importantes da distribuição dos dados que permitissem explicar similaridades ou dissimilaridades ou regularidades observadas entre as mensurações do fenômeno observado e seus locais de ocorrência. Para avaliação do papel das variáveis relativas ao manejo e das diferentes escalas da paisagem na determinação da diversidade, foi utilizada análise de correspondência e redundância (RDA).
5. RESULTADOS E DISCUSSÃO
5.1. Métricas das Diferentes Escalas da Paisagem e de Intensidade de Manejo
Esta primeira parte se dedicará a expor os dados de caracterização das diferentes escalas da paisagem agrícola das propriedades pertencentes às glebas A e B do assentamento São José da Boa Morte. Foram avaliados 14 parâmetros entre métricas da paisagem e de intensidade de manejo.
5.1.1. Escala do cultivo (0,5 ha)
O manejo adotado nos sistemas de cultivo (escala de 0,5 ha) avaliados apresentaram práticas de manejo similares alcançando valores bastante próximos dos parâmetros avaliados (Tab. 5), destacadamente no quesito agrotóxico (Agrtx). De maneira geral, os produtos comerciais utilizados possuem potencial de periculosidade ambiental de classe II (produto muito perigoso ao meio ambiente), havendo ao menos dois produtos com o PPA de classe I (produto altamente perigoso ao meio ambiente) utilizados no manejo de 3 das propriedades estudadas (A2, A4 e B3).
Quanto ao perfil da capina (Cap) adotada encontramos uma maior variabilidade. Na Gleba A, 60% das propriedades realizam apenas a capina mecânica ou associam a capina mecânica a um herbicida como forma de controle das espontâneas. Na Gleba B apenas em uma das propriedades a capina mecânica é adotada em associação com a capina química. Entre os herbicidas comerciais usados nas unidades produtivas avaliadas, 67% possuem o PPA de classe II (ex. Gramoxone; i.a.: Paraquat), sendo estes utilizados em 6 das 10 propriedades avaliadas ao total. Dos herbicidas utilizados na Gleba A, se caracterizam como não-seletivos os produtos comerciais de marca Gramocil e Roundup e seletivos para monocotiledôneas os herbicidas de marca Fusilade e Podium. Para a Gleba B, registrou-se a utilização apenas de herbicidas não-seletivos, das marcas comerciais Gramoxone e Roundup.
Quanto à intensidade de capinas realizadas (Int.cap) não houve diferença em ambas as glebas, tendo sido feita a média de 2 capinas durante o ciclo das culturas (CVC.c), que apresentou predominância de cultivos de ciclo médio na Gleba A, em especial a mandioca, e culturas de ciclo curto na Gleba B, sendo a batata-doce a cultura mais plantada.
Para o item “Solo”, foi observado que em maioria o manejo da adubação do solo na Gleba A se dá via adubo orgânico (valor 2), diferentemente da Gleba B em que há maior utilização de fertilizantes minerais (valor 1) por parte dos agricultores avaliados nessa área, em específico B2, B3 e B5.
Tabela 4. Parâmetros avaliados na escala do cultivo (0,5 ha), com média e respectivo desvio padrão, nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Onde: Agrtx (PPA dos agrotóxicos utilizados; adimensional), Cap (Tipo de capina utilizada; adimensional), Int.cap (Intensidade de capinas, adimensional), CVC.c (Ciclo da cultura; adimensional) e Solo (Tipo de adubação; adimensional).
|
Parâmetros |
Gleba A |
Gleba B |
|
Agrtx |
2,00 ± 1,64 |
2,00 ± 1,80 |
|
Cap |
3,09 ± 0,86 |
2,20 ± 0,50 |
|
Int.cap |
3,00 ± 0,84 |
3,60 ± 0,89 |
|
CVC.c |
2,00 ± 0,45 |
1,00 ± 0,55 |
|
Solo |
2,00 ± 0,50 |
1,00 ± 0,50 |
Fonte: Do próprio autor
5.1.2. Escala da unidade produtiva (4,0 ha)
Ambas as glebas apresentaram comportamento similar para todos os parâmetros propostos na escala da unidade produtiva (Tab. 6).
Para a avaliação do ciclo de vida das culturas (CVC.up) nota-se valores inferiores para a Gleba A, possivelmente pela predominância de culturas agrícolas anuais de ciclo médio (ex. mandioca) e curto (ex. batata-doce) que eram computadas com valores 2 e 1, respectivamente. Atualmente existem poucos pomares nas propriedades avaliadas nesta gleba, sendo presentes algumas poucas áreas com citros, que por ser uma cultura perene, conferiu valores maiores para estas propriedades, elevando o desvio padrão. Tal cenário se mostrou um pouco mais favorecido na Gleba B, que por conter propriedades com áreas destinadas ao cultivo de goiaba, aumentaram os valores deste índice.
Para o elemento arbóreo (EAup), ambas as glebas apresentaram características similares em suas propriedades, tendo a Gleba B uma média de 0,3 ha com cultivos de coco, eucalipto ou goiaba (EAup1). Entretanto este valor se dá por conta da propriedade B4 possuir em sua área um talhão de eucalipto com 1,7 ha, diluído na média das 6 propriedades. Já para áreas de vegetação secundária menores que 0,5 ha (EAup2) (ex. cerca viva com uma ou várias espécies ou vegetação de beira de rio), a Gleba A se caracteriza por possuir propriedades com média de 0,7 ha com este tipo de vegetação, sendo a propriedade A4 a que maior área possui, com 1,07 ha. A propriedade B1 é a que maior área destina a este parâmetro na Gleba B, com 0,63 ha, diferentemente da média de 0,03 ha para as demais propriedades desta gleba.
Não houve presença de áreas de fragmentos florestais na escala da unidade produtiva em ambas as glebas avaliadas.
Tabela 5. Valores médios e desvio padrão dos parâmetros avaliados na escala da unidade produtiva (4 ha) nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Onde: CVC.up (Ciclo da cultura; adimensional), EAup1 (Área com cultivo de coco, eucalipto e goiaba; ha), EAup2 (Área de vegetação secundária< 0,5 ha; ha) e AFF.up (Área de vegetação secundária > 0,5 ha; ha).
|
Parâmetros |
Gleba A |
Gleba B |
|
|
CVC.up |
1,50 ± 0,50 |
2,00 ± 0,16 |
|
|
EAup1 |
0,00 ± 0,00 |
0,27 ± 0,70 |
|
|
EAup2 |
0,70 ± 0,29 |
0,03 ± 0,27 |
|
|
AFF.up |
0,00 ± 0,00 |
0,00 ± 0,00 |
|
Fonte: Do próprio autor
5.1.3. Escala da paisagem (95,0 ha)
A caracterização descritiva através das métricas propostas para a escala da paisagem elucida a grande diferença na composição florestal das duas glebas avaliadas neste trabalho (Tab. 7).
Ao observarmos as médias da distância linear (Dist.p) entre o cultivo estabelecido no 0,5 ha e o fragmento florestal mais próximo, bem como a área do mesmo (Área.frag), nota-se que a Gleba A está imersa em uma paisagem com alta densidade florestal (x̄ = 96,52 ha ± 71,60), refletindo um mosaico de uso do solo em que ocorre de forma mais estreita as interações entre as fontes de biodiversidade próximas (x̄ = 228,69 ha ± 116,50) e os sistemas de cultivo. Este cenário se torna contrastante na caracterização da escala da paisagem da Gleba B, onde apenas há a existência de pequenos fragmentos isolados (x̄ = 12,35 ha ± 8,15) e distantes (x̄ = 640,69 m ± 386,49) das unidades produtivas.
Não existe diferença expressiva entre o item elemento arbóreo (EA.p) para os cultivos perenes (EAp1), entretanto para a vegetação secundária < 0,5 ha (EAp2) a Gleba A possui mais que o dobro da área deste item na Gleba B.
A área de fragmentos florestais presentes na escala da paisagem (AFF.p) foi o item com maior contraste entre as glebas avaliadas, com diferença de aproximadamente 10x na média das áreas estudadas em cada gleba (Gleba A = 20,27 ha ± 9,55; Gleba B = 2,28 ha ± 4,10).
Tabela 6. Parâmetros avaliados na escala da paisagem (100 ha), com médias e respectivos desvios padrão, nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Onde: Área.frag (Área do fragmento florestal mais próximo; ha), Dist.p (Distância do fragmento florestal mais próximo ao cultivo; m), EAp1 (Área com cultivo de coco, eucalipto e goiaba; ha), EAp2 (Área de vegetação secundária< 0,5 ha; ha), AFF.p (Área de vegetação secundária > 0,5 ha; ha).
|
Parâmetros |
Gleba A |
Gleba B |
|
|
Dist.p |
228,69 ± 116,50 |
640,69 ± 386,49 |
|
|
Área.frag |
96,52 ± 71,60 |
12,35 ± 8,15 |
|
|
EAp1 |
1,61 ± 3,61 |
2,81± 2,28 |
|
|
EAp2 |
7,69 ± 2,76 |
3,22 ± 1,19 |
|
|
AFF.p |
20,27 ± 9,55 |
2,28 ± 4,10 |
|
Fonte: Do próprio autor
Paisagens ricas em habitats naturais concedem um alto poder de resiliência para a matriz agrícola, dado a presença de fontes de propágulos, dispersores e alto grau de conectividade (TAMBOSI et al., 2013), como também o fornecimento de serviços ecossistêmicos essenciais para a agricultura (ZHANG et al., 2007; KREMEN & MILES, 2012).
Por sua vez, paisagens mais homogêneas e antropizadas nas diferentes escalas tendem a uma maior simplificação dos agroecossistemas, devido a perda de biodiversidade, levando a redução de serviços ecossistêmicos. Este ciclo de erosão de biodiversidade, perda de serviços ecossistêmicos e da resiliência, determina uma maior dependência do uso de insumos externos (TSCHARNTKE et al., 2005; KREMEN & MILES, 2012; ZHAO et al., 2015).
5.2. Avaliação da Fertilidade do Solo
A análise de fertilidade do solo (Tab. 8) quando expressada pela média das propriedades avaliadas em ambas as glebas, apresenta diferenças significativas entre as áreas para três dos oito elementos analisados e também para o pH, sendo estes elementos: Al, H+Al e P.
A diferença nos valores para os elementos citados pode se dar em função das diferentes práticas de manejo em relação à adubação, como também em função das características topográficas e de influência do lençol freático próximo à superfície do solo.
Tabela 7. Análise de fertilidade do solo das glebas A e B do assentamento São José da Boa Morte na escala de cultivo (0,5), representada pelas médias e desvios padrão das propriedades; (P<0,05).
|
Gleba |
C (%) |
Al (cmolc/d) |
Ca (cmolc/d) |
H+Al (cmolc/d) |
K (mg/L) |
Mg (cmolc/d) |
N (%) |
P (mg/L) |
pH |
|
A |
1,7 ± 1,4 a |
0,9 ± 1,2 a |
2,2 ± 1,3 a |
6,0 ± 4,8 a |
79,0 ± 65,9 a |
0,7 ± 0,5 a |
0,2 ± 0,1 a |
47,0 ± 16,5 a |
5,3 ± 0,9 a |
|
B |
2,2 ± 1,2 a |
1,5 ± 0,9 b |
1,6 ± 0,6 a |
8,8 ± 3,1 b |
83,2 ± 41,3 a |
0,6 ± 0,2 a |
0,2 ± 0,1 a |
31,1 ± 14,3 b |
4,6 ± 0,4 b |
Fonte: Do próprio autor
5.3. Descrição Geral da Comunidade de Espontâneas Amostradas in loco
Foram encontrados valores fitomassa total de espontâneas mais importantes na Gleba A quando comparada a Gleba B (37,5 t.ha-1 e 27,5 t.ha-1 respectivamente). Observando a composição da fitomassa tem-se que o percentual de dicotiledôneas encontrado na Gleba A foi maior que na Gleba B (31,5% e 23,6% respectivamente) (Tab. 9 e Fig. 12).
Tabela 8. Fitomassa (t.ha-1) da vegetação espontânea amostrada nas propriedades avaliadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ, classificada quanto ao seu morfotipo em: cyperáceas (Cyp); dicotiledôneas (Dicot); monocotiledôneas (Monocot).
|
Gleba A |
Gleba B |
||||||
|
Sistema |
Cyperáceas |
Dicot |
Monocot |
Sistema |
Cyperáceas |
Dicot |
Monocot |
|
1A |
0,8 |
2,3 |
2,6 |
1B |
4,0 |
2,6 |
0,6 |
|
2A |
4,0 |
1,6 |
0,0 |
2B |
1,4 |
2,1 |
2,9 |
|
3A |
4,4 |
0,6 |
1,3 |
3B |
1,1 |
0,1 |
0,3 |
|
4A |
2,1 |
2,3 |
1,4 |
4B |
2,0 |
1,3 |
0,7 |
|
5A |
1,5 |
5,1 |
2,5 |
5B |
2,8 |
0,5 |
2,3 |
|
6A |
3,1 |
0,2 |
1,9 |
6B |
2,5 |
0,2 |
0,0 |
|
x̄ |
2,6 |
2,0 |
1,6 |
2,3 |
0,9 |
0,7 |
|
|
DP |
1,4 |
1,7 |
1,0 |
1,1 |
1,0 |
1,2 |
|
|
CV |
0,55 |
0,90 |
0,58 |
0,46 |
1,13 |
1,71 |
|
Fonte: Do próprio autor
Para as Cyperáceas, os resultados encontrados para as duas glebas pode ser um reflexo das práticas de manejo do solo, aração frequente associada ao uso de herbicidas, que são hábitos comuns em muitas das propriedades avaliadas. A família Cyperaceae se caracteriza pelo seu estabelecimento rápido devido ao importante crescimento vegetativo e a produção de tubérculos, que na presença de condições ambientais favoráveis (temperatura elevada e intensa luminosidade) se intensifica (MILES et al., 1996). Tais características adaptativas, associadas ao manejo intensivo pela maior parte dos agricultores, permitem que essas plantas dominem a comunidade de espontâneas, simplificando os sistemas.
O coeficiente de variação (CV) para os grupos Monocotiledônea e Dicotiledônea se apresenta muito oscilante na Gleba B ao ser comparado aos valores para a Gleba A, podendo refletir a presença de plantas de ambos os grupos de forma mais constante na maior parte das propriedades avaliadas desta gleba. Estes valores podem inferir a existência de plantas espontâneas potenciais a serem caracterizadas mais especificamente quanto as suas funcionalidades e provisão de serviços ecossistêmicos.

Fonte: Do próprio autor
Figura 10. Fitomassa total (t.ha-1) da vegetação espontânea amostrada nas propriedades avaliadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu (RJ).


Fonte: Do próprio autor
Figura 11. Boxplot de distribuição da fitomassa (t.ha-1) da vegetação espontânea amostrada nas propriedades avaliadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu (RJ), classificada quanto ao seu morfotipo em: A = cyperáceas (Cyp); B = dicotiledôneas (Dicot); C = monocotiledôneas (Monocot). (Teste T; p<0,05).
Fonte: Do próprio autor
Figura 12. Distribuição percentual da fitomassa da vegetação espontânea amostrada nas propriedades avaliadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu (RJ), classificadas quanto ao seu morfotipo em: cyperáceas (vermelho), monocotiledôneas (laranja) e dicotiledôneas (azul).
5.4. Comunidade de Plantas Espontâneas do Banco de Sementes
Na comunidade de plantas espontâneas oriundas do banco de sementes foi registrado o total de 3997 indivíduos, distribuídos em 32 espécies de 18 famílias botânicas. Desse total, 1672 indivíduos foram obtidos de amostras originárias da Gleba A, distribuídos em 29 espécies de plantas emergidas do banco de sementes e 2325 indivíduos tiveram origem nas amostras da Gleba B, divididos em 30 espécies (Tab. 10).
As espécies Chamaesyce hirta e Oxalis sepium ocorreram apenas na Gleba A e as espécies Cuphea carthagenensis, Oldenlandia corymbosa e Sinapis arvensis apenas na Gleba B.
Destacaram-se as espécies Cyperus rotundus (21,50%), Digitaria ciliaris (18,40%) e Megathyrsus maximus (10,16%) como espécies de maior densidade relativa na Gleba A, representando 50,0% de todos os indivíduos emergidos na avaliação geral para essa área, e Cyperus iria (30,0%), Cyperus rotundus (15,0%), Ageratum conyzoides (5,0%) representam 51,0% da densidade relativa na Gleba B.
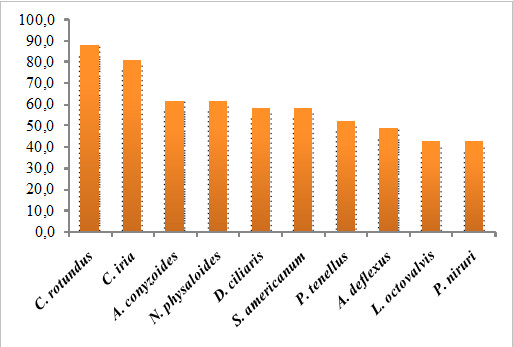
Para os cálculos de frequência relativa, em ambas as áreas a espécie Cyperus rotundus foi a mais frequente, estando em 87,0% das áreas, seguida por Digitaria ciliaris (77,4%), Magathyrsus maximus (58,0%) e pelas dicotiledôneas Nicandra physaloides (51,6%) e Phyllanthus tenellus (41,9%) como as mais frequentes na Gleba A (Fig. 13), e por Cyperus iria (80,65%), Ageratum conyzoides (61,7%) e Nicandra physaloides (61,7%) na Gleba B (Fig. 14).
.jpg)
Fonte: Do próprio autor
Figura 13. Gráfico com as dez espécies de maior frequência (%) na avaliação geral do banco de sementes de plantas espontâneas da Gleba A do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ.
Fonte: Do próprio autor
Figura 14. Gráfico com as dez espécies de maior frequência (%) na avaliação geral do banco de sementes de plantas espontâneas da Gleba B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ.
Entre as 10 espécies mais frequentes, expressaram-se cinco dicotiledôneas na Gleba A e sete na Gleba B, incluindo Amaranthus deflexus (48,4%) nessa área, entrando em acordo com Vasileiadis et al. (2007) que indica que as espécies de Amaranthus sp. tendem a sempre ser uma das mais expressivas no banco de sementes do solo em áreas de atividade agrícola.
Monquero & Christoffoleti (2005) mencionam que a quantidade de propágulos no banco de sementes de plantas espontâneas é, comparativamente, maior em áreas agrícolas do que em áreas não-agrícolas com baixo distúrbio ambiental. Esta tendência consiste em estratégia das plantas espontâneas, principalmente as mais ruderais, de produzir grandes quantidades de sementes em ambientes que apresentem alto distúrbio. No banco de sementes são encontradas muitas espécies, porém as poucas espécies dominantes compreendem em torno de 70 a 90% do total.
A predominância de Cyperus spp. como gênero mais frequente no banco de sementes do solo das propriedades avaliadas reflete a importância desse gênero como um dos mais importantes do mundo entre as plantas espontâneas devido à sua grande capacidade de reprodução vegetativa e disseminação, o que torna bastante difícil o seu controle. A movimentação do solo no sistema convencional de manejo é a principal forma de disseminação desta espécie em todo o mundo. Isto ocorre porque, com o revolvimento do solo pelos implementos de discos ou de hastes, ocorre a fragmentação e deslocamento dos tubérculos da planta, os quais originam inúmeros propágulos a cada operação de preparo (CORDEIRO et al., 2006).
5.4.1. Análise de agrupamento de espécies (cluster)
As medidas de distância de uma maneira geral podem ser definidas como medidas de similaridade e dissimilaridade, na qual a primeira é para definir o grau de semelhança entre as instâncias e realizam o agrupamento de acordo com a sua coesão, e a segunda de acordo com as diferenças dos atributos das instâncias (WITTEN e FRANK, 2005).
.jpg)
Fonte: Do próprio autor
Figura 15. Análise de agrupamento das propriedades agrícolas avaliadas com base na frequência relativa das espécies emergidas do banco de sementes do solo. A e B referem-se às propriedades das glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Coeficiente de correlação cofenética = 0,905.
A construção do cluster (Fig. 15) objetivou proporcionar a melhor visualização da similaridade das propriedades avaliadas em cada gleba em função da composição florística. O coeficiente de correlação cofenética de 0,905 expressa forte aderência dos dados para o modelo proposto. Nota-se a formação de três grupos, sob domínio de dois grupos principais. No primeiro, estão as propriedades A6, A5 e B4, e no segundo as demais propriedades, sendo que A1, A2 e A3 e B2, B5 e B6 possuem maior similaridade, da mesma forma que B3, A4 e B1 se encontram um terceiro grupo mais similar.
Diferentes práticas de manejo e intensidade do sistema produtivo, vetores ambientais das escalas de maior abrangência (unidade produtiva e paisagem) associados às síndromes de dispersão das diferentes espécies podem esclarecer a organização surgida para as propriedades, haja vista a heterogeneidade desses diferentes elementos em cada sistema avaliado. Os mesmos serão mais bem elucidados nos itens a seguir.
5.4.2. Agrupamento das espécies espontâneas pelo NMDS
O escalonamento multidimensional não-paramétrico (NMDS) expressou um STRESS (STandard REsiduals Sum of Squares ou Padrão Residual da Soma dos Quadrados) de 0,198 sendo considerado razoável para representar uma medida do quanto as posições de objetos em uma configuração tridimensional desviam-se das distâncias originais ou similaridades após o escalonamento.
.jpg)
Fonte: Do próprio autor
Figura 16. Escalonamento multidimensional não-paramétrico (NMDS), feito com base na frequência relativa de indivíduos do banco de sementes de plantas espontâneas e sua distribuição nas propriedades. Locais precedidos da letra A e B representam as propriedades situadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Ag.con (Ageratum conyzoides); Ama.def (Amaranthus deflexus); Bid.pil (Bidens pilosa); Cha.hi. (Chamaescyce hirta); Com.ben (Commelina benghalensis); Cuph.cat (Cuphea carthagenensis); Cyp.iria (Cyperus iria); Cyp.rot (Cyperus rotundus); Dig.cil (Digitaria ciliaris); Dio.sap (Diodia saponariifolia); Ele.ind (Eleusine indica); Emil.fo (Emilia fosbergii); Emil.son (Emilia sonchifolia); Euph.het (Euphorbia heterophylla); Gal.parv (Galinsoga parviflora); Gna.spi (Gnaphalium spicatum); Lep.vir (Lepidium virginicum); Lud.oct (Ludwigia octovalvis); Mega.ma (Megathyrsus maximus); Nic.phy (Nicandra physaloides); Old.cor (Oldenlandia corimbosa); Ox.cor (Oxalis corniculata); Ox.lat (Oxalis latifolia); Ox.sep (Oxalis sepium); Phy.nir (Phyllanthus niruri); Phy.ten (Phyllanthus tenellus); Port.ol (Portulaca oleracea); Sid.rom (Sida rhombifolia); Sin.arv (Sinapis arvensis); Sol.am (Solanum americanum); Ste.vert (Stemodia verticillata); Indet (espécie não identificada).
A análise apresenta uma organização das propriedades quanto às espécies de espontâneas emergidas do banco de sementes do solo, ao observar no eixo NMDS1 uma distribuição em dois grupos evidentes, separando as propriedades das duas glebas estudadas, sendo A4 o ponto mais distinto dessa organização.
Houve uma maior concentração de espécies de maior plasticidade ao centro do gráfico, aproximando as propriedades A1, A2, B2, B4, B5, nas quais estas foram mais frequentes.
O eixo NMDS2 demonstra uma distribuição vertical mais heterogênea para as propriedades da Gleba B do que para as da Gleba A, que possui com maior similaridade as espécies C. hirta, C. iria, E. heterophylla, L. virginicum, O. latifolia, O. sepium e S. verticillata.
As propriedades B1, B3 e B4 se agruparam em função das espécies B. pilosa, D. ciliaris, C. benghalensis, C. rotundus, L. octovalvis, P. niruri e S. rhombifolia.
Regensburgerd et al. (2008) destacaram algumas estratégias de dispersão de sementes, sendo algumas delas: anemocoria (sementes transportadas pelo vento), endozoocoria (sementes passam pelo trato digestivo dos animais para serem dispersas), epizoocoria (transportadas na pelagem dos animais) e autocoria (disseminação própria da semente pelo próprio fruto). No entanto, em áreas com influência antrópica, como é o caso de propriedades agrícolas, se torna importante adicionar outras formas de dispersão como a hidrocoria (disseminação pela água) sendo presente em cultivos irrigados e a antropocoria, já que o homem é um importante agente dispersor de sementes de plantas espontâneas, seja diretamente, pela utilização de sementes ou mudas contaminadas, seja indiretamente como, por exemplo, através do uso de implementos ou sacarias que não foram suficientemente limpas e que podem distribuir sementes de uma área a outra (DEUBER, 1992).
Espécies pertencentes à família Asteraceae, que possuem predominantemente dispersão anemocórica, se apresentaram distribuídas por todas as propriedades avaliadas, tendo as espécies A. conyzoides e Emilia sonchifolia como as de maior disseminação entre as áreas. Entretanto, esta síndrome de dispersão se aplica a maioria das especies emergidas nas avaliações realizadas nesse trabalho, apesar disso, espécies como B. pilosa, C. hirta, C. carthagenensis, G. spicatum, G. parviflora, O.corymbosa, Sida rhombifolia, entre outras, se localizaram de forma mais periférica, estando relacionadas a características mais específicas de algumas unidades produtivas.
Ao relacionar as práticas de controle de espontâneas realizada nas propriedades (ver parâmetro "Cap", item 4.1.1 - Escala do cultivo (0,5 ha)) com a distribuição das espécies, observa-se no gráfico que as espécies de reprodução predominantemente vegetativa estão próximas às propriedades da Gleba A, onde a maioria dos agricultores avaliados optam em realizar capina mecânica no lugar de capina química, podendo então propiciar uma maior individualização das estruturas vegetativas de espécies como D. saponariifolia, as Oxalidaceaes: O. corniculata, O. latifolia e O. sepium, e P. oleracea, aumentando a perpetuação destas em áreas mais localizadas.
5.5. Composição de Espontâneas em Grupos Funcionais e seus Serviços Ecossistêmicos Associados
A diversidade e composição de espontâneas em grupos funcionais pode desempenhar um papel importante no fornecimento e sustentabilidade de serviços ecossistêmicos, principalmente em paisagens agrícolas (POWER, 2010).
Ao observarmos a contribuição das espécies espontâneas quanto ao seu potencial de provimento de serviços à escala de cultivo, foi obtido de forma generalizada que 44% dos serviços são prestados pelo grupo funcional “Alimentação e Saúde Humana” (ASH), sendo 12 espécies com potencial para usos na alimentação humana (AH) e 25 para usos como plantas medicinais (UM).
Em seguida, o grupo referente ao potencial de contribuição para o controle de pragas e doenças (CPD) expressou 25% dos grupos funcionais, sendo 7 espécies para repelentes de pragas e doenças (RPD) e 15 com o potencial de atração de inimigos naturais (AIN). Segundo Muhammad et al. (2013), as plantas espontâneas acrescentam heterogeneidade fitomórfica e fornecem recursos alimentícios, auxiliando na manutenção de muitas populações de artrópodes incluindo espécies de fitófagos, desta forma, possibilita reduzir a infestação de pragas em plantas de interesse econômico. Também alguns predadores usam o estrato herbáceo como hospedeiro alternativo em diferentes fases do ciclo de vida, tanto para abrigo quanto para se alimentar de insetos-praga (NEWTON, 2004; ABBAS et al., 2012).
Alguns trabalhos em ecologia de paisagens agrícolas sugerem que sistemas produtivos com maior diversidade de comunidades florais respondem positivamente às alterações produzidas por adversidades ambientais (TSCHARNTKE et al., 2005). Em nosso estudo, 17 espécies compõem o grupo com características que apoiam o estímulo à polinização (EP), sendo este responsável por 18% dos serviços ambientais prestados (Gleba A = 15; Gleba B = 16). Ashiq et al. (2003) relatou cerca de 50 espécies em seu trabalho, dos quais apenas 15 espécies forneciam recursos florais ao sistema. A aplicação de agrotóxicos, fertilizantes e herbicidas podem ser considerados elementos responsáveis por esta lacuna funcional (SIDDIQUI, 2005).
Quanto ao aumento da qualidade do solo, (AQS) representa 14% dos serviços prestados, sendo 3 espécies para o suporte a aspectos físicos (EsS), e 9 que contribuem para a melhoria da fertilidade do solo (FS), estabelecendo condições mais aptas à posterior colonização por outras espécies ou para a melhoria de áreas destinadas à cultivos (Tab. 10).
De acordo com Costa & Mitja (2009), uma maior diversidade de espécies de plantas implica em diferentes sistemas radiculares, o que pode contribuir para uma melhor estruturação e aeração do solo, favorecendo o desenvolvimento microbiano e, consequentemente, a fauna do solo.
Tabela 9. Número total de espécies por grupos funcionais e seus respectivos serviços ecossistêmicos associados nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ, sendo: Alimentação Humana (AH); Uso Medicinal (UM); Estruturação do Solo (EsS); Fertilidade do Solo (FS); Repelente de Pragas e Doenças (RPD); Atração de Inimigos Naturais (AIN); Estímulo a polinização (EP).
|
Áreas |
Alimentação e Saúde Humana |
Aumento da Qualidade do Solo |
Controle de Pragas e Doenças |
Estímulo à Polinização (EP) |
|||
|
(ASH) |
(AQS) |
(CPD) |
|||||
|
AH |
UM |
EsS |
FS |
RPD |
AIN |
EP |
|
|
Gleba A |
12 |
25 |
3 |
9 |
7 |
15 |
15 |
|
Gleba B |
13 |
27 |
3 |
9 |
6 |
16 |
16 |
Fonte: Do próprio autor
A tabela a seguir (Tab. 12) expressa a riqueza de espécies e abundância de indivíduos por serviços e grupos funcionais prestados nas diferentes propriedades e glebas avaliadas.
Tabela 10. Riqueza (S), abundância total (AB) e número de espécies por grupos funcionais e seus respectivos serviços ecossistêmicos associados, nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ, sendo: Alimentação Humana (AH); Uso Medicinal (UM); Estruturação do Solo (EsS); Fertilidade do Solo (FS); Repelente de Pragas e Doenças (RPD); Atração de Inimigos Naturais (AIN); Estímulo à Polinização (EP).
|
Áreas |
S |
AB |
Alimentação e Saúde Humana |
Aumento da Qualidade do Solo |
Controle de Pragas e Doenças |
Estímulo à Polinização (EP) |
|||
|
(ASH) |
(AQS) |
(CPD) |
|||||||
|
AH |
UM |
EsS |
FS |
RPD |
AIN |
EP |
|||
|
A1 |
11 |
196 |
4 |
10 |
1 |
1 |
5 |
6 |
4 |
|
A2 |
17 |
419 |
5 |
16 |
1 |
5 |
5 |
10 |
11 |
|
A3 |
20 |
355 |
9 |
18 |
6 |
3 |
6 |
11 |
10 |
|
A4 |
16 |
267 |
8 |
15 |
2 |
6 |
5 |
10 |
8 |
|
A5 |
13 |
355 |
6 |
12 |
1 |
5 |
6 |
7 |
8 |
|
A6 |
11 |
79 |
4 |
10 |
2 |
3 |
4 |
5 |
6 |
|
B1 |
14 |
445 |
8 |
13 |
1 |
4 |
6 |
8 |
7 |
|
B2 |
17 |
315 |
7 |
16 |
1 |
8 |
5 |
9 |
8 |
|
B3 |
16 |
185 |
8 |
15 |
2 |
5 |
6 |
11 |
8 |
|
B4 |
13 |
461 |
5 |
11 |
1 |
4 |
3 |
7 |
6 |
|
B5 |
18 |
457 |
9 |
17 |
0 |
7 |
5 |
8 |
11 |
|
B6 |
20 |
459 |
9 |
18 |
1 |
7 |
5 |
8 |
11 |
Fonte: Do próprio autor
Ao observar a tabela acima, nota-se como os diferentes serviços estão sendo suportados nas diferentes propriedades avaliadas, e quais desses possuem maior estabilidade em função da maior quantidade de espécies com papéis funcionais similares, já que a redundância de espécies é considerada necessária para garantir a resiliência dos ecossistemas à perturbação (WALKER, 1995).
Ao observar o desvio padrão das diferentes propriedades para os serviços avaliados, foi observado que para os serviços inclusos no grupo ‘Aumento da Qualidade do Solo’ (AQS) há uma desviação de ±0,5 para ‘EsS’ e de ±2,2 para ‘FS’ nas propriedades da gleba A e de ±1,2 e ±0,8 para os mesmo serviços na gleba B, sendo este o único grupo que apresentou maiores valores para essa avaliação. Para os demais serviços não houve diferenças aparentes tampouco quando comparado os valores entre as glebas A e B.
5.6. Análise de Redundância (RDA) para a Associação dos Serviços Ecossistêmicos à Intensidade de Manejo e aos Componentes da Paisagem nas Escalas Avaliadas
A análise de redundância (RDA, na sigla em inglês) foi utilizada para detectar o quanto da mudança na variável resposta (matriz da composição de serviços ecossistêmicos) pode ser explicado pelas variáveis ambientais e de intensidade de manejo nas diferentes escalas avaliadas, determinando se, e quanto, as variáveis são correlacionadas.
5.6.1. RDA - Escala de cultivo (0,5 ha)
A RDA para a avaliação da escala de cultivo (0,5 ha) apresentou significância estatística (p = 0,042) para os componentes avaliados a um rigor de 5%. Os dados serão discutidos a seguir.
.jpg)
Fonte: Do próprio autor
Figura 17. Análise de redundância (RDA) para os parâmetros de avaliação da intensidade de manejo e dos serviços ecossistêmicos. Locais precedidos da letra A e B representam as propriedades situadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. Agrtx (PPA dos agrotóxicos utilizados; adimensional); CVC.c (Ciclo de vida da cultura no 0,5 ha; adimensional) Int.cap (Relação entre tipo e intensidade de capina, adimensional); Solo (Tipo de adubação do solo; adimensional); AH (Alimentação Humana); UM (Uso Medicinal); EsS (Estruturação do Solo); FS (Fertilidade do Solo); RPD (Repelente de Pragas e Doenças); AIN (Atração a Inimigos Naturais); EP (Estímulo à Polinização); p = 0,042.
Abaixo, segue a contribuição dos eixos da RDA para os parâmetros avaliados (Tabelas 13, 14 e 15).
Tabela 11. Importância do RDA1 e RDA2 para a relação Escala do Cultivo (0,5 ha)/ Serviços Ecossistêmicos.
|
Componentes |
RDA1 |
RDA2 |
|
Autovalores |
0,1380 |
0,1069 |
|
Proporção explicada |
0,4509 |
0,3494 |
|
Proporção cumulativa |
0,8003 |
|
Fonte: Do próprio autor
Tabela 12. Contribuição dos eixos RDA1 e RDA2 na avaliação dos serviços ecossistêmicos para a escala do cultivo (0,5 ha).
|
Serviços ecossistêmicos |
RDA1 |
RDA2 |
|
AH |
0,3428 |
-0,1195 |
|
UM |
0,1297 |
0,2892 |
|
EsS |
0,1334 |
0,0052 |
|
FS |
0,3201 |
-0,2956 |
|
RPD |
-0,3654 |
0,0583 |
|
AIN |
0,0430 |
0,4789 |
|
EP |
0,4570 |
0,2146 |
Fonte: Do próprio autor
Tabela 13. Contribuição dos eixos RDA1 e RDA2 na avaliação da intensidade de manejo na escala do cultivo (0,5 ha).
|
Intensidade de Manejo |
RDA1 |
RDA2 |
|
CVC.c |
-0,0635 |
0,5040 |
|
Int.cap. |
-0,8448 |
0,5103 |
|
Agrtx |
-0,7842 |
-0,5156 |
|
Solo |
-0,1811 |
0,0846 |
Fonte: Do próprio autor
Os serviços mais influenciados pelos parâmetros da intensidade de cultivo foram EP, FS, sendo os vetores: intensidade de capina (Int.cap.) e agrotóxicos (Agrtx), os mais expressivos desses parâmetros, podendo determinar significativamente a composição do banco de sementes do solo ao longo do tempo, pois, como já posto, o controle da vegetação espontânea via aplicação de herbicidas se caracteriza como a prática agrícola que exerce maior influência negativa sobre a riqueza e abundância de espécies, sendo um dos principais filtros para a comunidade de espontâneas (BOOTH & SWANTON, 2002).
Apesar de pouca interferência no eixo RDA1, o vetor referente ao ciclo das culturas na escala de cultivo (CVC.c) teve importância para o posicionamento das propriedades A2, A3 e B3, e principalmente ao serviço relacionado à atração de inimigos naturais (AIN). Agroecossistemas podem ser considerados áreas de abrigo à biodiversidade, apresentando locais para nidificação e forrageamento de inimigos naturais, especialmente as culturas com ciclo mais longo bem como plantas de porte arbóreo se mostram mais amigáveis à conservação de insetos prestadores de serviços ambientais, como abelhas e vespas (GEMMILL-HERREN & OCHIENG, 2008; SHACKELFORD et al., 2013).
Desta forma, compreender os efeitos de diferentes práticas agrícolas para o banco de sementes e riqueza de espécies pode ajudar a desenvolver estratégias bem-sucedidas de manejo de espontâneas, evitando concorrência com as plantas de interesse econômico e auxiliando da diversificação da paisagem (JOSÉ-MARIA & SANS, 2011).
A maioria das propriedades da Gleba B se apresentaram agrupadas na parte inferior do gráfico, sendo B2, B4 e B6 no quadrante inferior esquerdo e B5 no quadrante inferior direto, próximo às propriedades A5 e A6, todas sob influência do vetor que representa o peso do componente ‘Agrtx’, muito provavelmente em função de que a maioria dos agricultores responsáveis pelo manejo destas áreas utilizam agrotóxicos menos intensivamente. Entretanto, seu distanciamento dos vetores ‘Int.cap’ e ‘CVC.c’, indica que o maior uso de herbicidas e a produção de culturas agrícolas de ciclo curto influenciam negativamente na prestação de serviços ecossistêmicos.
Os resultados obtidos para essa escala corroboram com os estudos de Booth & Swanton, (2002), Armengot et al. (2011) e José-Maria & Sans (2011), que dialogam sobre o impacto da intensificação de práticas de manejo em unidades produtivas sobre, principalmente, a riqueza e densidade de espécies espontâneas, a partir do momento que nossos dados sinalizam que sistemas produtivos com menor intensidade de capina, ou seja, que lançam mão de menor utilização de herbicidas e maior espaço entre aplicações, beneficiam a emergência de espécies espontâneas funcionalmente importantes.
5.6.2. RDA - Escala da unidade produtiva (4,0 ha)
Para os parâmetros propostos em nosso estudo para a escala da unidade produtiva (4,0 ha), o RDA não apontou a existência de correlação entre tais componentes, obtendo um valor de p = 0,715, superior ao rigor de 5%.
.jpg)
Fonte: Do próprio autor
Figura 18. Análise de redundância (RDA) para os parâmetros de avaliação da escala da unidade produtiva (4,0 ha) e dos serviços ecossistêmicos. Locais precedidos da letra A e B representam as propriedades situadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. CVC.up (Ciclo da cultura; adimensional), EAup1 (Área com cultivo de coco, eucalipto e goiaba; ha), EAup2 (Área de vegetação secundária < 0,5 ha; ha); AH (Alimentação Humana); UM (Uso Medicinal); EsS (Estruturação do Solo; (FS (Fertilidade do Solo); RPD (Repelente de Pragas e Doenças); AIN (Atração a Inimigos Naturais); EP (Estímulo à Polinização); p = 0,715.
A seguir, as tabelas com a contribuição dos eixos da RDA para os parâmetros avaliados (Tabelas 16, 17 e 18).
Tabela 14. Importância do RDA1 e RDA2 para a relação dos componentes da escala da unidade produtiva (4,0 ha) e os serviços ecossistêmicos.
|
Componentes |
RDA1 |
RDA2 |
|
Autovalores |
0,0836 |
0,0376 |
|
Proporção explicada |
0,1432 |
0,0645 |
|
Proporção cumulativa |
0,2078 |
|
Fonte: Do próprio autor
Tabela 15. Contribuição dos eixos RDA1 e RDA2 na avaliação dos serviços ecossistêmicos na escala da unidade produtiva (4,0 ha).
|
Serviços ecossistêmicos |
RDA1 |
RDA2 |
|
AH |
0,0056 |
0,0938 |
|
UM |
-0,3729 |
-0,0067 |
|
EsS |
0,0070 |
-0,3462 |
|
FS |
0,2393 |
0,0887 |
|
RPD |
0,0817 |
-0,0743 |
|
AIN |
-0,3097 |
-0,0586 |
|
EP |
-0,2528 |
0,1342 |
Fonte: Do próprio autor
Tabela 16. Contribuição dos eixos RDA1 e RDA2 na avaliação dos componentes da escala da unidade produtiva (4,0 ha).
|
Componentes und. produtiva |
RDA1 |
RDA2 |
|
CVC.up |
-0,0551 |
0,1093 |
|
EAup1 |
0,9561 |
0,0361 |
|
EAup2 |
-0,4581 |
-0,8857 |
Fonte: Do próprio autor
Embora não haja significância estatística para os componentes avaliados, observa-se uma forte expressão do vetor relacionado à presença de áreas de vegetação secundária menor que 0,5 ha (EAup2), principalmente no eixo RDA2, porém, não sendo possível realizar inferências quanto a importância deste tipo de elemento para o banco de sementes de espontâneas e na determinação da prestação de serviços na escala avaliada.
5.6.3. RDA - Escala da paisagem (95,0 ha)
Para um rigor de 5%, a RDA em questão obteve significância estatística (p = 0,033), expressando correlação positiva entre os componentes propostos para a escala da paisagem e os serviços ecossistêmicos avaliados, sendo estes discutidos a seguir.
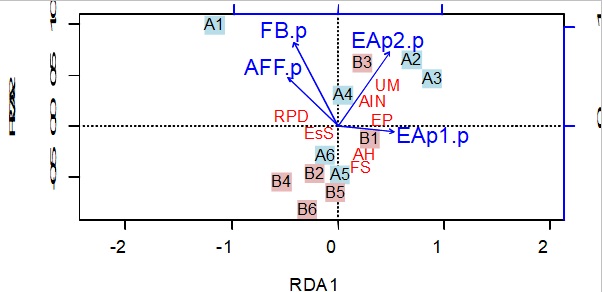
Fonte: Do próprio autor
Figura 19. Análise de redundância (RDA) para os parâmetros de avaliação da escala da paisagem (95,0 ha) e dos serviços ecossistêmicos; Locais precedidos da letra A e B representam as propriedades situadas nas glebas A e B do assentamento São José da Boa Morte, Cachoeiras de Macacu, RJ. FB.p (Fonte de Biodiversidade = Relação distância/área do fragmento florestal mais próximo ao cultivo), EAp1 (Área com cultivo de coco, eucalipto e goiaba; ha), EAp2 (Área de vegetação secundária < 0,5 ha; ha), AFF.p (Área de vegetação secundária > 0,5 ha; ha); AH (Alimentação Humana); UM (Uso Medicinal); EsS (Estruturação do Solo); FS (Fertilidade do Solo); RPD (Repelente de Pragas e Doenças); AIN (Atração de Inimigos Naturais); EP (Estímulo à Polinização); p = 0,033.
Abaixo, segue a contribuição dos eixos da RDA para os parâmetros avaliados (Tabelas 19, 20 e 21).
Tabela 17. Importância do RDA1 e RDA2 para a relação dos componentes da escala da paisagem (95 ha) e os serviços ecossistêmicos.
|
Componentes |
RDA1 |
RDA2 |
|
Autovalores |
0,1877 |
0,0976 |
|
Proporção explicada |
0,5898 |
0,3066 |
|
Proporção cumulativa |
0,8964 |
|
Fonte: Do próprio autor
Tabela 18. Contribuição dos eixos RDA1 e RDA2 na avaliação dos serviços ecossistêmicos na escala da paisagem (95 ha).
|
Serviços ecossistêmicos |
RDA1 |
RDA2 |
|
AH |
0,3016 |
-0,2479 |
|
UM |
0,4058 |
0,3731 |
|
EsS |
-0,0419 |
0,0498 |
|
FS |
0,2753 |
-0,3478 |
|
RPD |
-0,4520 |
0,1285 |
|
AIN |
0,2677 |
0,2881 |
|
EP |
0,4536 |
0,0046 |
Fonte: Do próprio autor
Tabela 19. Contribuição dos eixos RDA1 e RDA2 na avaliação dos componentes da escala da paisagem (95,0 ha).
|
Componentes da paisagem |
RDA1 |
RDA2 |
|
EA.p1 |
0,4993 |
0,7516 |
|
EA.p2 |
0,5411 |
-0,0590 |
|
AFF.p |
-0,4793 |
0,4940 |
|
FB.p |
-0,4243 |
0,8455 |
Fonte: Do próprio autor
A análise reflete a presença de dois grupos distintos pelo eixo RDA2, alocando as propriedades da Gleba A nos quadrantes superiores, próximas aos vetores associados aos componentes da paisagem, e outro grupo nos quadrantes inferiores, concentrando a maioria das propriedades da Gleba B e outras duas da Gleba A (A5 e A6).
Todos os vetores exibiram importância para o sistema, principalmente ao que relaciona a proximidade a fragmentos florestais mais densos, representando fontes de biodiversidade (FB.p), como também a presença de áreas de vegetação secundária menores que 0,5 ha (EAup2), que influenciou diretamente a provisão de serviços relacionados ao potencial de usos medicinais (UM) e atração de inimigos naturais (AIN), aproximando as propriedades A2, A3 e B3. Autores como Gabriel et al. (2005) e Roschewitz et al. (2005) apontam que paisagens mais complexas podem beneficiar a diversificação das espécies de espontâneas em áreas de cultivo, comparado a paisagens simplificadas, uma vez que o primeiro oferece um cenário em que existe uma maior quantidade de habitats, atuando como fonte de propágulos para o enriquecimento e manutenção do banco de sementes em áreas agrícolas.
O eixo RDA1 exibe a influência que os vetores: áreas com cultivo de coco, eucalipto e goiaba, e as de vegetação secundária (EA.p1 e EA.p2, respectivamente) tiveram sobre os serviços: uso medicinal (UM), estímulo à polinização (EP) e alimentação humana (AH), apontando a importância de se manter em propriedades agrícolas, pequenos bosques, cercas vivas, cultivos em aléias seja com utilização de arbóreas para fornecimento de matéria orgânica, madeira ou frutíferas. Entre os dois componentes, o que está relacionado às áreas de coco, eucalipto e goiaba obteve maior peso em ambos os eixos, respaldando a hipótese de que a presença do elemento arbóreo pode determinar a composição funcional de espontâneas, sendo importante tal informação, pois em áreas de baixa densidade florestal, representado em nosso trabalho pela Gleba B, tais áreas de cultivo podem ser a única forma de promover melhores condições para a emergência de plantas espontâneas menos competitivas, por alterações no microhabitat, impulsionando a diversificação do estrato herbáceo, que pode representar uma das poucas zonas de amortecimento e fonte de alimentos para inimigos naturais, polinizadores, entre outros organismos em agroecossistemas imersos em paisagens simplificadas e com reduzidas fontes de biodiversidade, impulsionando minimamente a biodiversidade na escala de cultivo.
Já os vetores relacionados à presença de áreas de fragmentos florestais e fontes de biodiversidade (AFF.p e FB.p) tiveram, para esse eixo, maior peso para o serviço relacionado a plantas repelentes de pragas e doenças (RPD). Esses mesmos vetores imprimiram forte influência para as características da propriedade A1, que se posicionou no gráfico distanciada de todas as demais.
O serviço de estruturação do solo (EsS) não se mostrou afetado pelos componentes avaliados para essa escala.
6. CONCLUSÕES
Nossos resultados adicionam aos de estudos anteriores que destacam a importância de se considerar as espécies e diversidades funcionais simultaneamente na compreensão das respostas dos serviços ecossistêmicos à intensificação das diferentes escalas da paisagem agrícola, pois expressamos que:
-
As práticas de manejo realizadas na escala de cultivo, em especial o tipo e intensidade de capina e as fontes de biodiversidade na escala da paisagem determinam a composição do banco de sementes de plantas espontâneas.
-
A prestação de serviços ecossistêmicos pela vegetação espontânea pode ser ampliada com a inserção do elemento arbóreo na unidade produtiva, sejam como pequenos bosques ou pomares.
-
Plantas espontâneas, quando diversas funcionalmente, representam uma das poucas alternativas para amenizar o declínio da biodiversidade em agroecossistemas pobres de áreas fonte.
7. CONSIDERAÇÕES FINAIS
Em um cenário onde a agricultura é apontada como uma das pressões mais significativas à erosão da biodiversidade, principalmente em ecossistemas tropicais, readequações nas atividades exercidas em matrizes agrícolas intensificadas devem receber destaque na atenção de gestores públicos e responsáveis pela assistência técnica e extensão rural em áreas de atividade agropecuária próximas a fragmentos florestais.
Estudos anteriores a este, realizados na mesma localidade, apontaram que a provisão dos serviços ecossistêmicos de vespas e abelhas (ex. controle biológico e polinização) são garantidos em unidades produtivas próximas a fontes de biodiversidade e que utilizam menos insumos nas práticas de manejo adotadas.
Sendo assim, a construção de politicas públicas e o investimento de recursos que visem identificar elementos prestadores de serviços ecossistêmicos para a agricultura, diversificando a paisagem agrícola e ofertando potenciais para usos econômicos se faz essencial para promover um modelo de agricultura aderente às demandas ambientais atuais.
O presente trabalho visou contribuir com informações que direcionem para tal caminho, compreendendo alguns dos vetores de influência na prestação de serviços por parte das plantas espontâneas em agroecossistemas, elucidando a importância das mesmas na construção e manutenção da agrobiodiversidade. Entretanto, a disponibilidade de informações ainda é muito escassa e pouco condizente com o perfil agrícola e ambiental das diferentes regiões do Brasil, sendo então de grande importância o estímulo à produção do conhecimento nas abordagens aqui expostas.
8. REFERÊNCIAS BIBLIOGRÁFICAS
ALTIERI, M. A. Linking ecologists and traditional farmers in the search for sustainable agriculture. Frontiers in Ecology and the Environment, v. 2, n. 1, p. 35-42, 2004.
ALVES, R. C. Efeito da interação do padrão de cultivo e das diferentes escalas de uma paisagem agrícola sobre a diversidade de vespas e abelhas. Seropédica, 57p. Monografia de conclusão de curso (Graduação em Engenharia Florestal). Instituto de Florestas, Universidade Federal Rural do Rio de Janeiro, 2013.
ANDREASEN, C.; STREIBIG, J. C. Evaluation of changes in weed flora in arable fields of Nordic countries–based on Danish long‐term surveys. Weed Research, v. 51, n. 3, p. 214-226, 2011.
ANGELSEN, A.; KAIMOWITZ, D. (Ed.). Agricultural technologies and tropical deforestation. CABi, 2001.
ARMENGOT, L. et al. Landscape and land-use effects on weed flora in Mediterranean cereal fields. Agriculture, ecosystems & environment, v. 142, n. 3, p. 311-317, 2011.
ARMENGOT, L. et al. Weed harrowing in organically grown cereal crops avoids yield losses without reducing weed diversity. Agronomy for sustainable development, v. 33, n. 2, p. 405-411, 2013.
ASHIQ, M.; NAYYAR, M. M.; AHMED, J. Weed Control Handbook for Pakistan. Directorate of Agronomy. Ayub Agricultural Research Institute, Faisalabad, p. 11-184, 2003.
ASSESSMENT, Millennium Ecosystem. Ecosystems and human well-being: desertification synthesis. World Resources Institute, 2005.
BAILEY, D. et al. Effects of habitat amount and isolation on biodiversity in fragmented traditional orchards. Journal of Applied Ecology, v. 47, p. 1003–1013, 2010.
BÀRBERI, P., SILVESTRI, N., BONARI, E.Weed communities of winter as influenced by imput level and rotation. Weed Res., v.37, n.5, p.301-13, 1997.
BÀRBERI, P. et al. Functional biodiversity in the agricultural landscape: relationships between weeds and arthropod fauna. Weed Research, v. 50, n. 5, p. 388-401, 2010.
BENTON, T. G. et al. Linking agricultural practice to insect and bird populations: a historical study over three decades. Journal of Applied Ecology, v. 39, n. 4, p. 673–687, 2002.
BIANCHI, F. J. J. A.; BOOIJ, C. J. H.; TSCHARNTKE, T. Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proceedings of the Royal Society of London B: Biological Sciences, v. 273, n. 1595, p. 1715-1727, 2006.
BIANCHI, F. J. J. A. et al. Opportunities and limitations for functional agrobiodiversity in the European context. Environmental Science & Policy, v. 27, p. 223-231, 2013
BLACKSHAW, R. E.; MOLNAR, L. J.; JANZEN, H. H. Nitrogen fertilizer timing and application method affect weed growth and competition with spring wheat. Weed Science, v. 52, n. 4, p. 614-622, 2004
BLACKSHAW, R. E.; MOLNAR, L. J.; LARNEY, F. J. Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada. Crop Protection, v. 24, n. 11, p. 971-980, 2005.
BOOTH, B. D.; SWANTON, C. J. Assembly theory applied to weed communities. 2009.
BRUSSAARD, L.; DE RUITER, P. C.; BROWN, G. G. Soil biodiversity for agricultural sustainability. Agriculture, ecosystems & environment, v. 121, n. 3, p. 233-244, 2007.
BRUSSAARD, L. et al. Reconciling biodiversity conservation and food security: scientific challenges for a new agriculture. Current opinion in Environmental sustainability, v. 2, n. 1, p. 34-42, 2010.
CARMONA, R. Banco de sementes e estabelecimento de plantas daninhas em agroecossistemas. Planta Daninha, Viçosa, v. 13, n. 1, p.3-9, 1995.
CHAPLIN‐KRAMER, R. et al. A meta‐analysis of crop pest and natural enemy response to landscape complexity. Ecology letters, v. 14, n. 9, p. 922-932, 2011.
CHRISTOFFOLETI, P. J.; LÓPEZ-OVEJERO, R. F.; Resistência das plantas daninhas a herbicidas: definições, bases e situação no Brasil e no mundo. Aspectos de resistência de plantas daninhas a herbicidas, v. 3, p. 9-34, 2008.
CLIMATE DATA, 2013. Disponível em: <http://pt.climate-data.org/location/33704>, Acesso em: 25 Mai. 2015.
CINCOTTA, R. P.; WISNEWSKI, J.; ENGELMAN, R.. Human population in the biodiversity hotspots. Nature, v. 404, n. 6781, p. 990-992, 2000.
Conservation International do Brasil. Grandes Regiões Naturais: as últimas áreas silvestres da Terra. Brasil: Conservation National, 1998.
COSTA, J. R.; MITJA, D.; FONTES, J. R. A. Bancos de sementes de plantas daninhas em cultivos de mandioca na Amazônia Central = Weed seed banks in Cassava cultivations in Central Amazon. Planta Daninha, v. 27, n. 4, p. 665-671, 2009.
CORDEIRO, L.A.M.; REIS, M.S.; AGNES, E.L.; CECON, P.R. Efeito do Plantio Direto no Controle de Tiririca (Cyperus rotundus L.) e Outras Plantas Daninhas na Cultura do Milho. Revista Brasileira de Herbicidas, Passo Fundo-RS, n. 1, p 1-9. 2006.
DALE, V. H.; POLASKY, S. Measures of the effects of agricultural practices on ecosystem services. Ecological economics, v. 64, n. 2, p. 286-296, 2007.
DEUBER, R. Ciência das plantas daninhas: fundamentos. Jaboticabal: FUNEP, v. 1, 431 p. 1992.
DI TOMASO, J. M. Approaches for improving crop competitiveness through the manipulation of fertilization strategies. Weed Science, p. 491-497, 1995.
DORÉ, T., MAKOWSKI, D., MALÉZIEUX, E., MUNIER-JOLAIN, N., TCHAMITCHIAN, M. & TITTONELL, P. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. European Journal of Agronomy, 34, 197-210. 2011.
ESTRADA, A.; COATES-ESTRADA, R.. Dung beetles in continuous forest, forest fragments and in an agricultural mosaic habitat island at Los Tuxtlas, Mexico. Biodiversity & Conservation, v. 11, n. 11, p. 1903-1918, 2002.
EWERS, R. M.; DIDHAM, R. K. Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews, v. 81, p. 117-142, 2006.
EVANS, D. M. et al. Seeds in farmland food-webs: resource importance, distribution and the impacts of farm management. Biological conservation, v. 144, n. 12, p. 2941-2950, 2011.
FAO. Agricultural Biodiversity, Multifunctional Character of Agriculture and Land Conference, Background Paper 1. 1999a.
FAHRIG, L. Rethinking patch size and isolation effects: the habitat amount hypothesis. Journal of Biogeography, v. 40, n. 9, p. 1649-1663, 2013.
FIDALGO, E. C. C.; PEDREIRA, B. C. C. G.; ABREU, M. B.; MOURA, I. B.; GODOY, M. D. P. Uso e cobertura da terra na bacia hidrográfica do rio Guapi-Macacu. Rio de Janeiro: Embrapa Solos (Embrapa Solos. Documentos, 105). 2008.
FISCHER, J. et al. Should agricultural policies encourage land sparing or wildlife-friendly farming?. Frontiers in Ecology and the Environment, v. 6, n. 7, p. 380-385, 2008.
FLYNN, D. FB et al. Loss of functional diversity under land use intensification across multiple taxa. Ecology letters, v. 12, n. 1, p. 22-33, 2009.
FOLEY, J. A. et al. Solutions for a cultivated planet. Nature, v. 478, n. 7369, p. 337-342, 2011.
FRANKE AC, L. L., VAN DER BURG W.J. & VAN OVERBEEK L. The role of arable weed seeds for agroecosystem functioning. Weed Research 49, 131-141. 2009.
GABRIEL, Doreen; THIES, Carsten; TSCHARNTKE, Teja. Local diversity of arable weeds increases with landscape complexity. Perspectives in Plant Ecology, Evolution and Systematics, v. 7, n. 2, p. 85-93, 2005.
GABRIEL, D. et. al. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecology Letters, v. 13, p. 858-869, 2010.
GARDNER, J. B.; DRINKWATER, L. E. The fate of nitrogen in grain cropping systems: a meta-analysis of 15N field experiments.Ecological Applications, v. 19, n. 8, p. 2167-2184, 2009.
GEMMILL-HERREN, B.; OCHIENG, A. O. Role of native bees and natural habitats in eggplant (Solanum melongena) pollination in Kenya.Agriculture, Ecosystems & Environment, v. 127, n. 1, p. 31-36, 2008.
GREEN, R. E. et al. Farming and the fate of wild nature. Science, v. 307, n. 5709, p. 550-555, 2005.
GUERRERO, I. et al. Influence of agricultural factors on weed, carabid and bird richness in a Mediterranean cereal cropping system. Agriculture, ecosystems & environment, v. 138, n. 1, p. 103-108, 2010.
GUERRERO, I. et al. Non-linear responses of functional diversity and redundancy to agricultural intensification at the field scale in Mediterranean arable plant communities. Agriculture, Ecosystems & Environment, v. 195, p. 36-43, 2014.
HAUGGAARD-NIELSEN, H.; JENSEN, E. S. Facilitative root interactions in intercrops. Plant and Soil, v. 274, n. 1-2, p. 237-250, 2005.
HAWES, C. et al. Arable plant communities as indicators of farming practice.Agriculture, ecosystems & environment, v. 138, n. 1, p. 17-26, 2010.
HENDRICKX, F. et. al. How landscape structure, land-use intensity and habitat diversity affect components of total arthropod diversity in agricultural landscapes. Journal of Applied Ecology, v. 44, p. 340-351, 2007.
HOLLAND, J. M. et al. A review of invertebrates and seed‐bearing plants as food for farmland birds in Europe. Annals of Applied Biology, v. 148, n. 1, p. 49-71, 2006.
HOOPER, D. U. et al. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological monographs, v. 75, n. 1, p. 3-35, 2005.
IBAMA - Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis. Consolidação de dados fornecidos pelas empresas registrantes de produtos técnicos, agrotóxicos e afins, conforme art. 41 do Decreto n° 4.074/2002. 2014.
IBAMA – Instituto Brasileiro de Meio Ambiente e dos Recursos Naturais. Disponível em: <www.ibama.gov.br/qualidade-ambiental/avaliacao-do-potencial-de-periculosidade-ambiental-ppa/pagina-5> Acesso em: 18/05/2015.
JACKSON, L. E.; PASCUAL, U.; HODGKIN, T. Utilizing and conserving agrobiodiversity in agricultural landscapes. Agriculture, ecosystems & environment, v. 121, n. 3, p. 196-210, 2007.
JONGMANS, A. G. et al. Soil structure and characteristics of organic matter in two orchards differing in earthworm activity. Applied Soil Ecology, v. 24, n. 3, p. 219-232, 2003.
GROSS, K. L. A comparison of methods for estimating seed numbers in the soil. The Journal of Ecology, p. 1079-1093, 1990.
KINUPP, V. F. Plantas alimentícias não-convencionais da região metropolitana de Porto Alegre, RS. 2007.
KORASAKI, V. et al. Conservation value of alternative land-use systems for dung beetles in Amazon: valuing traditional farming practices.Biodiversity and conservation, v. 22, n. 6-7, p. 1485-1499, 2013.
KOTTEK, M. et. al. World Map of the Köppen-Geiger climate classification updated. Meteorologische Zeitschrift, v. 15, n. 3, p. 259-263, 2006.
KREMEN, C.; WILLIAMS, N. M.; THORP, R. W. Crop pollination from native bees at risk from agricultural intensification. Proceedings of the National Academy of Sciences, v. 99, n. 26, p. 16812-16816, 2002.
KREMEN, C.; ILES, A.; BACON, C.. Diversified farming systems: an agroecological, systems-based alternative to modern industrial agriculture. 2012.
LACERDA, A. L. de S.; VICTORIA FILHO, R.; MENDONÇA, C. G. Levantamento do banco de sementes em dois sistemas de manejo de solo irrigados por pivô central. Planta Daninha, Viçosa, v. 23, p. 1-7, 2005.
LETOURNEAU, D. K. et al. Effects of natural enemy biodiversity on the suppression of arthropod herbivores in terrestrial ecosystems. Annual Review of Ecology, Evolution, and Systematics, v. 40, p. 573-592, 2009.
LIN, B. B. Agroforestry management as an adaptive strategy against potential microclimate extremes in coffee agriculture. Agricultural and Forest Meteorology, v. 144, n. 1, p. 85-94, 2007.
LOPES, P. FM; SILVANO, R. AM; BEGOSSI, A. Extractive and Sustainable Development Reserves in Brazil: resilient alternatives to fisheries?. Journal of Environmental Planning and Management, v. 54, n. 4, p. 421-443, 2011.
LOREAU, M.; MOUQUET, N.; GONZALEZ, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proceedings of the National Academy of Sciences, v. 100, n. 22, p. 12765-12770, 2003.
MACHADO, V. M. et al. Evaluation of the seed bank in a campestre cerrado area under recovery. Planta Daninha, v. 31, n. 2, p. 303-312, 2013.
MACLEAN, R.N. Impact of gliricidia spectabalis hedgerows on weeds and insect pests of upland rice. Agriculture, Ecosystems and Environment, v.94, p. 275-288. 2003.
MAHN, E. Changes in the structure of weed communities affected by agro-chemicals: what role does nitrogen play?. Ecological Bulletins, p. 71-73, 1988.
MAYFIELD, M. M.; LEVINE, J. M. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecology letters, v. 13, n. 9, p. 1085-1093, 2010.
MONQUERO, P. A.; CHRISTOFFOLETI, P. J. Banco de sementes de plantas daninhas e herbicidas como fator de seleção. Bragantia, v. 64, n. 2, p. 203-209, 2005.
MORANDIN, L. A.; WINSTON, M. L. Wild bee abundance and seed production in conventional, organic, and genetically modified canola. Ecological applications, v. 15, n. 3, p. 871-881, 2005.
MOREIRA, E. F.; BOSCOLO, D.; VIANA, B. F. Spatial heterogeneity regulates plant-pollinator networks across multiple landscape scales. 2015.
MUHAMMAD, S. et al. Impact of organic fertilizer, humic acid and sea weed extract on wheat production in Pothowar region of Pakistan. Pak. J. Agri. Sci, v. 50, n. 4, p. 677-681, 2013.
MULVANY, P.; BERGER, R. Biodiversidad Agrıcola: Cuando los agricultores mantienen la red de la vida. Entendiendo la Biodiversidad Agrıcola, v. 1, p. 14-16, 2003.
MURPHY, C. E.; LEMERLE, D. Continuous cropping systems and weed selection. Euphytica, v. 148, n. 1-2, p. 61-73, 2006.
NEWTON, I. The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions. Ibis, v. 146, n. 4, p. 579-600, 2004
NIE, J. et al. Weed community composition after 26 years of fertilization of late rice. Weed science, v. 57, n. 3, p. 256-260, 2009.
ORŁOWSKI, G.; CZARNECKA, J. Winter diet of reed bunting Emberiza schoeniclus in fallow and stubble fields. Agriculture, ecosystems & environment, v. 118, n. 1, p. 244-248, 2007.
PASCUAL, U.; PERRINGS, C. Developing incentives and economic mechanisms for in situ biodiversity conservation in agricultural landscapes.Agriculture, Ecosystems & Environment, v. 121, n. 3, p. 256-268, 2007.
PEARCE, D. W.; MORAN, D. The economic value of biodiversity. Earthscan, 1994.
PERFECTO, I.; VANDERMEER, J. The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proceedings of the National Academy of Sciences, v. 107, n. 13, p. 5786-5791, 2010.
PETCHEY, O. L.; GASTON, K. J. Functional diversity (FD), species richness and community composition. Ecology Letters, v. 5, n. 3, p. 402-411, 2002.
PHALAN, B. et al. Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science, v. 333, n. 6047, p. 1289-1291, 2011.
PIMM, S. L. The balance of nature?: Ecological issues in the conservation of species and communities. University of Chicago Press, 1991.
POWER, A. G. Ecosystem services and agriculture: tradeoffs and synergies. Philosophical transactions of the royal society B: biological sciences, v. 365, n. 1554, p. 2959-2971, 2010.
R Core Team (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. Disponível em: (http://www.R-project.org/). Acesso em: 20 Jun 2015.
REGENSBURGER, B.; COMIN, J.J; AUMOND, J. J. Integração de técnicas de solo, plantas e animais para recuperar áreas degradadas. Ciência Rural, v.38, n.6, set, 2008.
RICKETTS, T. H. et al. Landscape effects on crop pollination services: are there general patterns?. Ecology letters, v. 11, n. 5, p. 499-515, 2008
RIEDER, A. et al. Classes de potencial de periculosidade ambiental (ppa) dos pesticidas receitados em municípios do pantanal norte, Mato Grosso (MT), Brasil, no biênio 1999-2000. in: Simposio sobre recursos naturais e sócio-econômicos do Pantanal. 2004.
ROSCHEWITZ, I. et al. The effects of landscape complexity on arable weed species diversity in organic and conventional farming. Journal of Applied Ecology, v. 42, n. 5, p. 873-882, 2005.
SCARIOT, A. Land sparing or land sharing: the missing link. Frontiers in Ecology and the Environment, v. 11, n. 4, p. 177-178, 2013.
SCHERR, S. J.; MCNEELY, J. A. Biodiversity conservation and agricultural sustainability: towards a new paradigm of ‘ecoagriculture’landscapes. Philosophical Transactions of the Royal Society B: Biological Sciences, v. 363, n. 1491, p. 477-494, 2008.
SEA – Secretaria de Estado do Ambiente. Disponível em: <http://www.rj.gov.br/web/sea/exibeconteudo?article-id=164974> Acessado em 18 de Mai. de 2015.
SHACKELFORD, N. et al. Primed for change: developing ecological restoration for the 21st century. Restoration Ecology, v. 21, n. 3, p. 297-304, 2013.
SIDDIQI, M. J. I. Studies on the biodiversity of invertebrates in the wheat Triticum aestivum farm agroecosystems of Punjab, Pakistan. 2005. Tese de Doutorado. Ph. D. Thesis. Department of Zoology & Fisheries, University of Agriculture, Faisalabad, Pakistan.
SLADE, E. M.; MANN, D. J.; LEWIS, O. T. Biodiversity and ecosystem function of tropical forest dung beetles under contrasting logging regimes. Biological Conservation, v. 144, n. 1, p. 166-174, 2011.
SMITH, Richard G. Timing of tillage is an important filter on the assembly of weed communities. Weed Science, v. 54, n. 4, p. 705-712, 2006.
STORKEY, J.; CUSSANS, J. W. Reconciling the conservation of in-field biodiversity with crop production using a simulation model of weed growth and competition. Agriculture, ecosystems & environment, v. 122, n. 2, p. 173-182, 2007.
STORKEY, J. et al. Using functional traits to quantify the value of plant communities to invertebrate ecosystem service providers in arable landscapes. Journal of ecology, v. 101, n. 1, p. 38-46, 2013.
TAMBOSI, L. R. et al. A framework to optimize biodiversity restoration efforts based on habitat amount and landscape connectivity. Restoration ecology, v. 22, n. 2, p. 169-177, 2014.
TANG, L. et al. Impact of fertilizing pattern on the biodiversity of a weed community and wheat growth. 2014.
TILMAN, D. et al. Diversity and productivity in a long-term grassland experiment. Science, v. 294, n. 5543, p. 843-845, 2001.
TOMICH, T. P. et al. Agroecology: a review from a global-change perspective. Annual Review of Environment and Resources, v. 36, p. 193-222, 2011.
TSCHARNTKE, T. et. al. Landscape perspectives on agricultural intensification and biodiversity – ecosystem service management. Ecology Letters, v. 8, p. 857-874, 2005.
TSCHARNTKE, T. et. al. Conservation biological control and enemy diversity on a landscape scale. Biological Control, v. 43, p. 294–309, 2007.
TSCHARNTKE, T. et. al. Landscape constraints on functional diversity of birds and insects in agroecosystems. Ecology, v. 89, p. 944–951, 2008.
ULBER, L. et al. An on‐farm approach to investigate the impact of diversified crop rotations on weed species richness and composition in winter wheat. Weed Research, v. 49, n. 5, p. 534-543, 2009.
ULBER, L.; STEINMANN, Horst-Henning; KLIMEK, Sebastian. Using selective herbicides to manage beneficial and rare weed species in winter wheat. Journal of Plant Diseases and Protection, p. 233-239, 2010.
UZÊDA, M. C.; Construção coletiva de princípios agroecológicos voltados à consolidação de paisagens sustentáveis no assentamento São José da Boa Morte (Cachoeiras de Macacu/ RJ). Rio de Janeiro. Embrapa Agrobiologia. 2012.
VANDERMEER, J. et al. Breakfast of biodiversity: the truth about rain forest destruction. Institute for Food and Development Policy, 1995.
VIVIAN, R. et al. Weed seed dormancy as a survival mechanism: brief review. Planta Daninha, v. 26, n. 3, p. 695-706, 2008.
WALKER, B. Conserving biological diversity through ecosystem resilience. Conservation biology, p. 747-752, 1995.
WITTEN, I. H., FRANK, E. Data Mining : practical machine learning tools and techniques. 2ª Edição – (2005). Morgan Kaufmann series in data management systems. ISBN: 0-12-088407-0
YIN, L.; CAI, Z.; ZHONG, W. Changes in weed composition of winter wheat crops due to long-term fertilization. Agriculture, ecosystems & environment, v. 107, n. 2, p. 181-186, 2005.
ZHANG, W. et. al. Ecosystem services and dis-services to agriculture. Ecological Economics, v. 64, p. 253-260, 2007.
ZHAO, Zi-H. et al. Effects of agricultural intensification on ability of natural enemies to control aphids. Scientific reports, v. 5, 2015.
Publicado por: Fernando Igne Rocha
O texto publicado foi encaminhado por um usuário do site por meio do canal colaborativo Monografias. Brasil Escola não se responsabiliza pelo conteúdo do artigo publicado, que é de total responsabilidade do autor . Para acessar os textos produzidos pelo site, acesse: https://www.brasilescola.com.























